

Microscopic and biochemical monitoring of endosomal trafficking and extracellular vesicle secretion in an endogenous in vivo model
- PMID: 36103151
- PMCID: PMC9473323
- DOI: 10.1002/jev2.12263
Microscopic and biochemical monitoring of endosomal trafficking and extracellular vesicle secretion in an endogenous in vivo model
Abstract
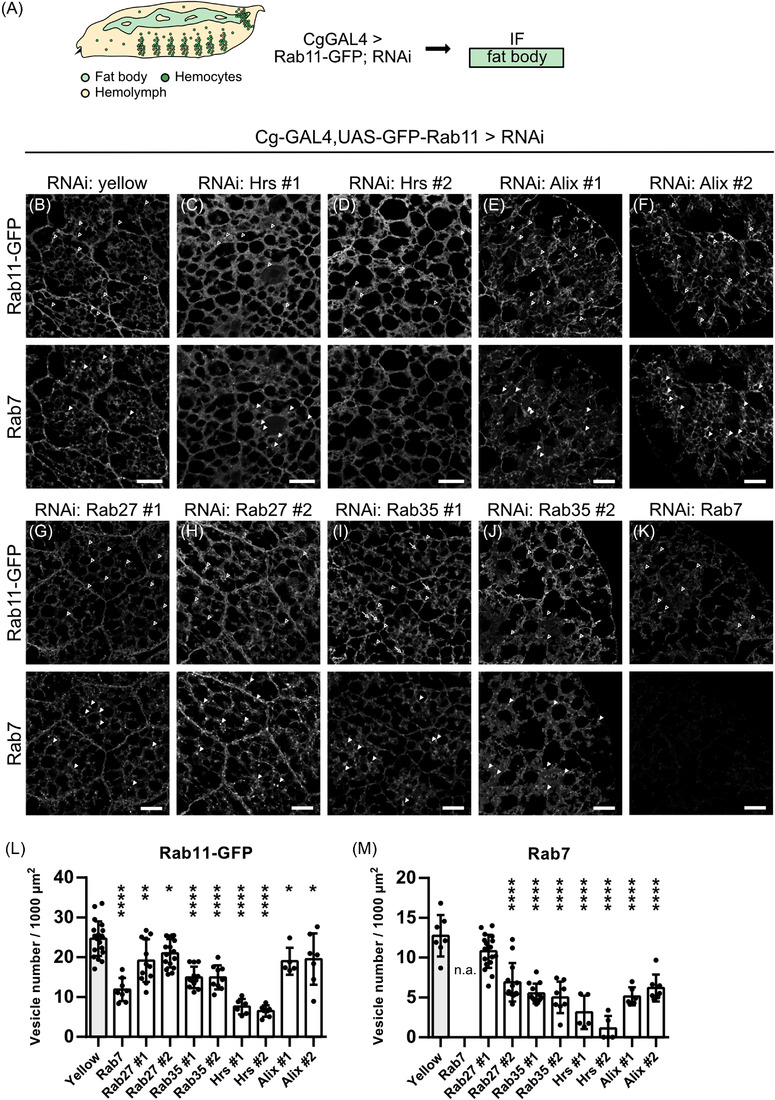
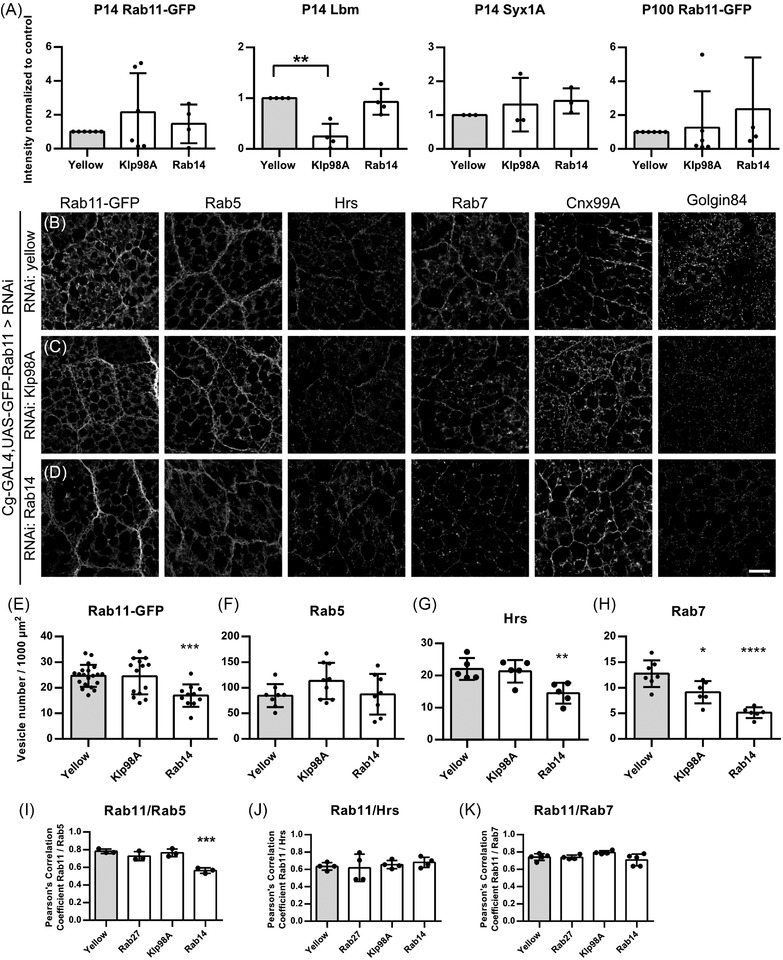
Extracellular vesicle (EV) secretion enables cell-cell communication in multicellular organisms. During development, EV secretion and the specific loading of signalling factors in EVs contributes to organ development and tissue differentiation. Here, we present an in vivo model to study EV secretion using the fat body and the haemolymph of the fruit fly, Drosophila melanogaster. The system makes use of tissue-specific EV labelling and is amenable to genetic modification by RNAi. This allows the unique combination of microscopic visualisation of EVs in different organs and quantitative biochemical purification to study how EVs are generated within the cells and which factors regulate their secretion in vivo. Characterisation of the system revealed that secretion of EVs from the fat body is mainly regulated by Rab11 and Rab35, highlighting the importance of recycling Rab GTPase family members for EV secretion. We furthermore discovered a so far unknown function of Rab14 along with the kinesin Klp98A in EV biogenesis and secretion.
Keywords: Drosophila; Rab11; Rab35; Tsp96F; exosomes; fat body; haemolymph; intercellular communication.
© 2022 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Conflict of interest statement
The authors have no competing financial interests.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
