

Synaptotagmin rings as high-sensitivity regulators of synaptic vesicle docking and fusion
- PMID: 36103579
- PMCID: PMC9499556
- DOI: 10.1073/pnas.2208337119
Synaptotagmin rings as high-sensitivity regulators of synaptic vesicle docking and fusion
Abstract
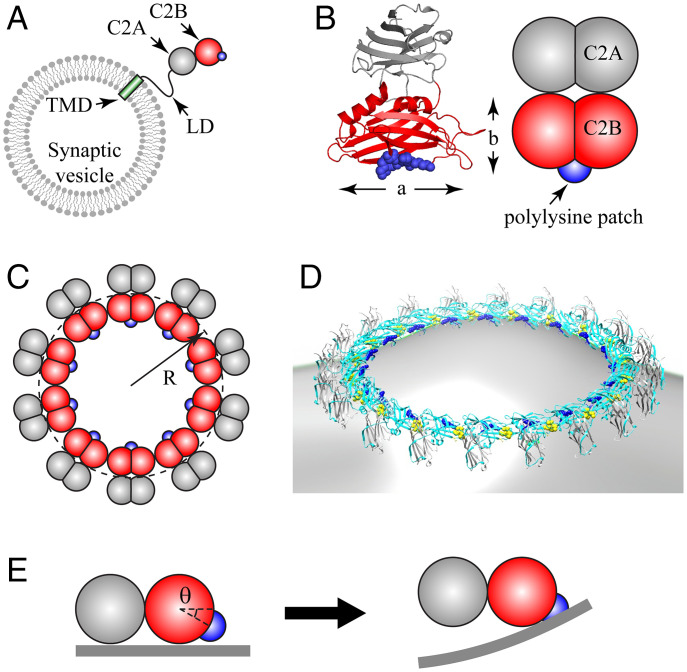
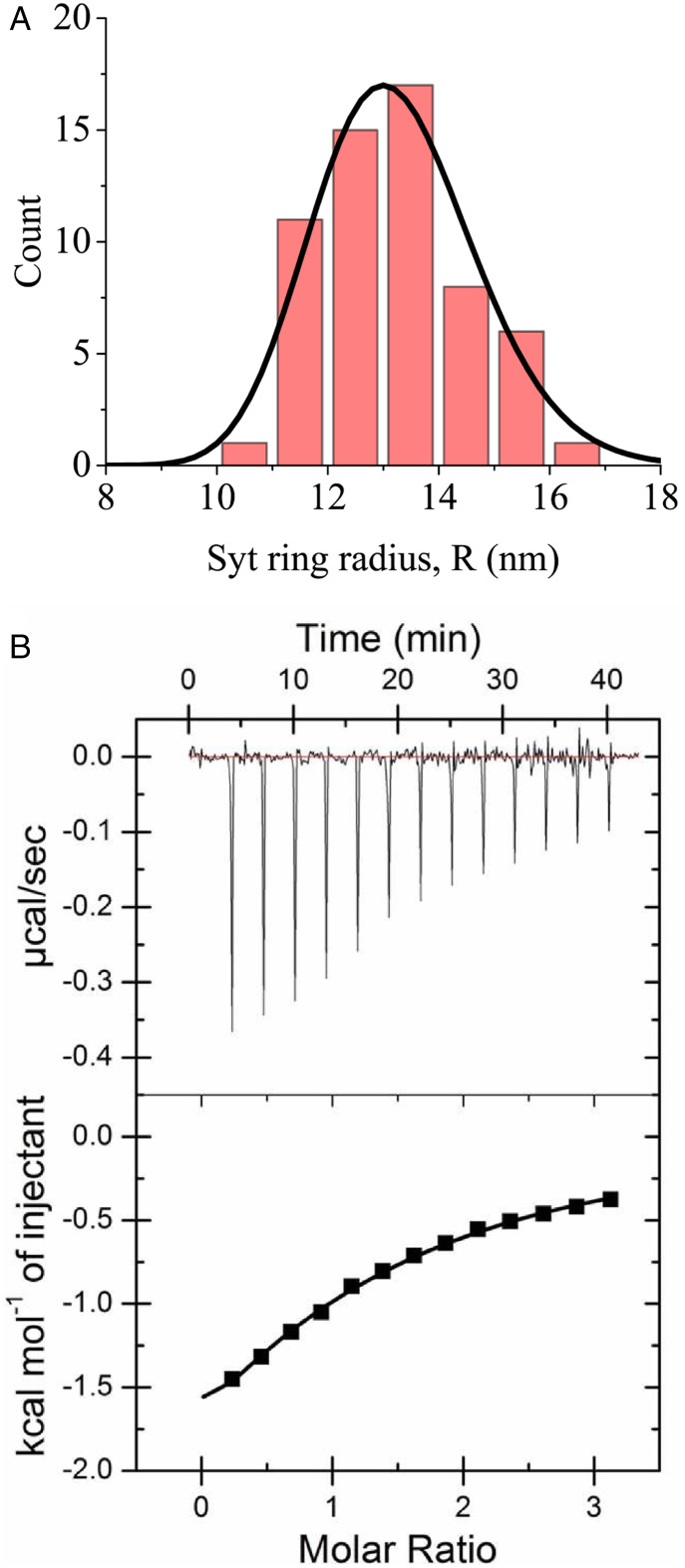
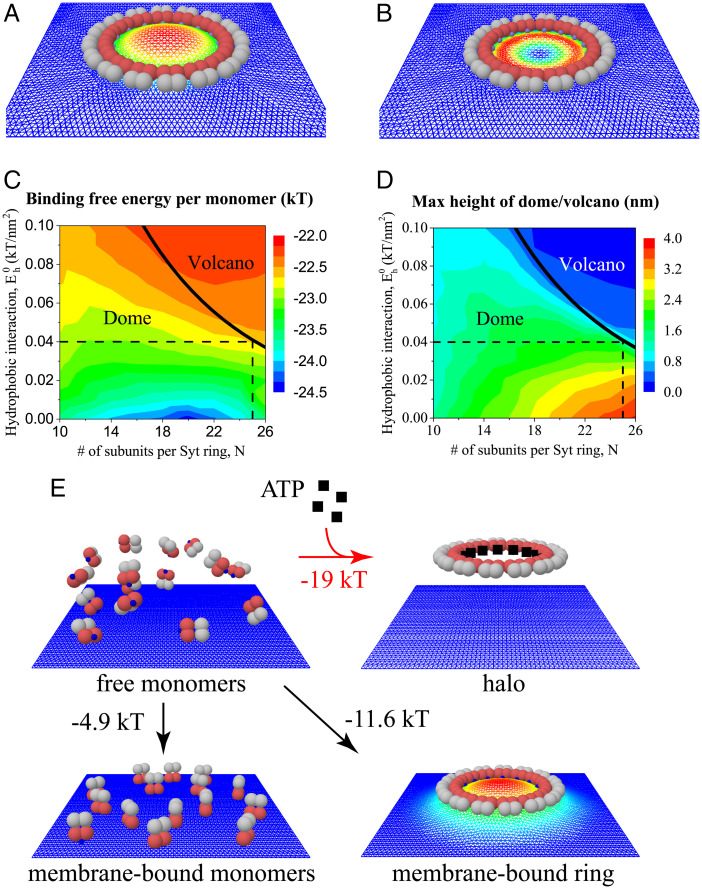
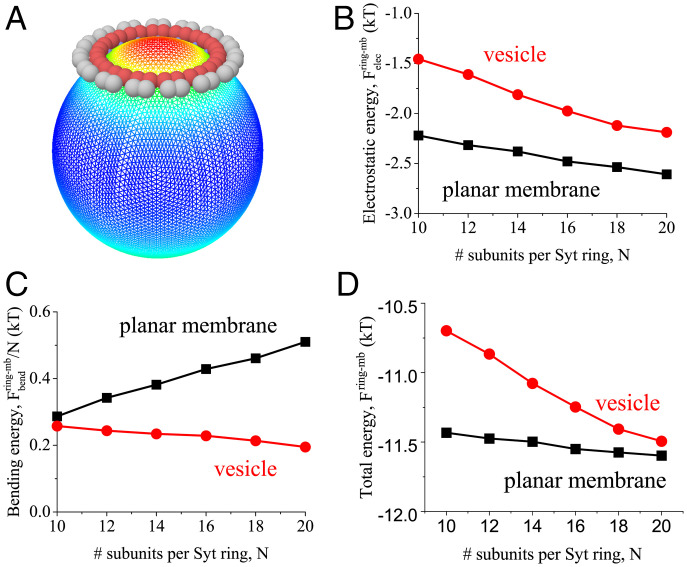
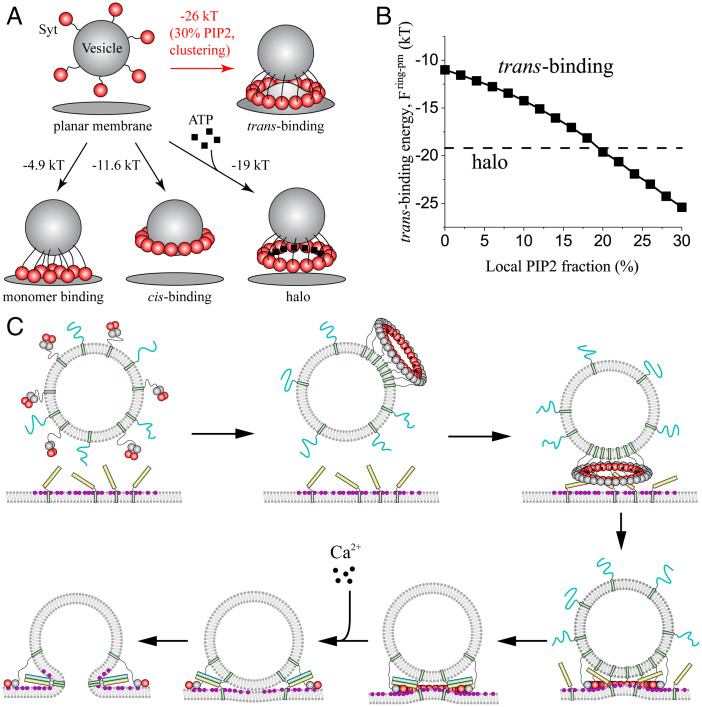
Synchronous release at neuronal synapses is accomplished by a machinery that senses calcium influx and fuses the synaptic vesicle and plasma membranes to release neurotransmitters. Previous studies suggested the calcium sensor synaptotagmin (Syt) is a facilitator of vesicle docking and both a facilitator and inhibitor of fusion. On phospholipid monolayers, the Syt C2AB domain spontaneously oligomerized into rings that are disassembled by Ca2+, suggesting Syt rings may clamp fusion as membrane-separating "washers" until Ca2+-mediated disassembly triggers fusion and release [J. Wang et al., Proc. Natl. Acad. Sci. U.S.A. 111, 13966-13971 (2014)].). Here, we combined mathematical modeling with experiment to measure the mechanical properties of Syt rings and to test this mechanism. Consistent with experimental results, the model quantitatively recapitulates observed Syt ring-induced dome and volcano shapes on phospholipid monolayers and predicts rings are stabilized by anionic phospholipid bilayers or bulk solution with ATP. The selected ring conformation is highly sensitive to membrane composition and bulk ATP levels, a property that may regulate vesicle docking and fusion in ATP-rich synaptic terminals. We find the Syt molecules hosted by a synaptic vesicle oligomerize into a halo, unbound from the vesicle, but in proximity to sufficiently phosphatidylinositol 4,5-bisphosphate (PIP2)-rich plasma membrane (PM) domains, the PM-bound trans Syt ring conformation is preferred. Thus, the Syt halo serves as landing gear for spatially directed docking at PIP2-rich sites that define the active zones of exocytotic release, positioning the Syt ring to clamp fusion and await calcium. Our results suggest the Syt ring is both a Ca2+-sensitive fusion clamp and a high-fidelity sensor for directed docking.
Keywords: membrane fusion; neurotransmitter release; synapse; synaptotagmin; vesicle docking.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Takamori S., et al. , Molecular anatomy of a trafficking organelle. Cell 127, 831–846 (2006). - PubMed
-
- Wilhelm B. G., et al. , Composition of isolated synaptic boutons reveals the amounts of vesicle trafficking proteins. Science 344, 1023–1028 (2014). - PubMed
-
- Chapman E. R., How does synaptotagmin trigger neurotransmitter release? Annu. Rev. Biochem. 77, 615–641 (2008). - PubMed
-
- Chapman E. R., Synaptotagmin: A Ca(2+) sensor that triggers exocytosis? Nat. Rev. Mol. Cell Biol. 3, 498–508 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
