

Loss of Epigenetic Regulation Disrupts Lineage Integrity, Induces Aberrant Alveogenesis, and Promotes Breast Cancer
- PMID: 36108220
- PMCID: PMC9812400
- DOI: 10.1158/2159-8290.CD-21-0865
Loss of Epigenetic Regulation Disrupts Lineage Integrity, Induces Aberrant Alveogenesis, and Promotes Breast Cancer
Abstract
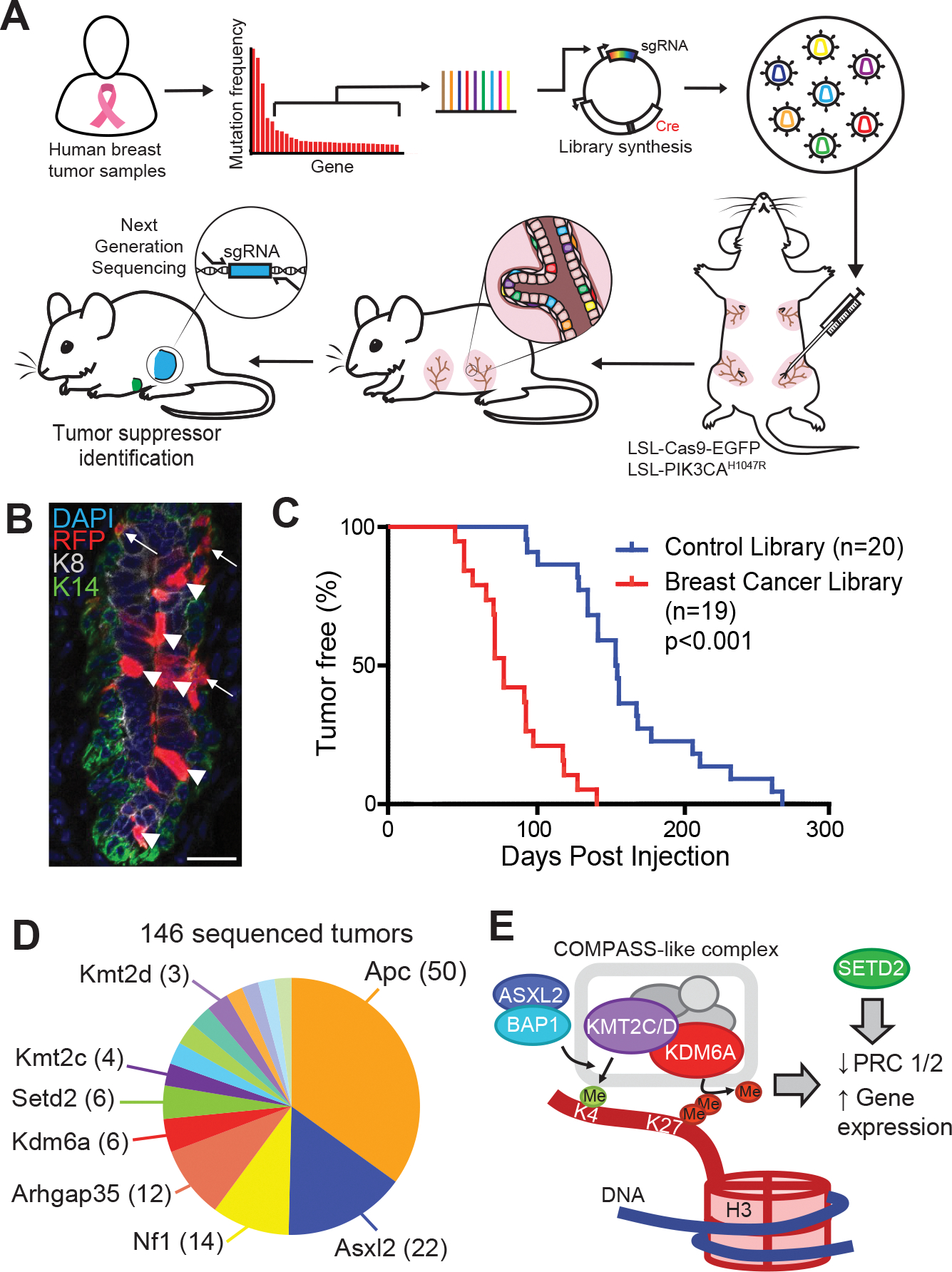
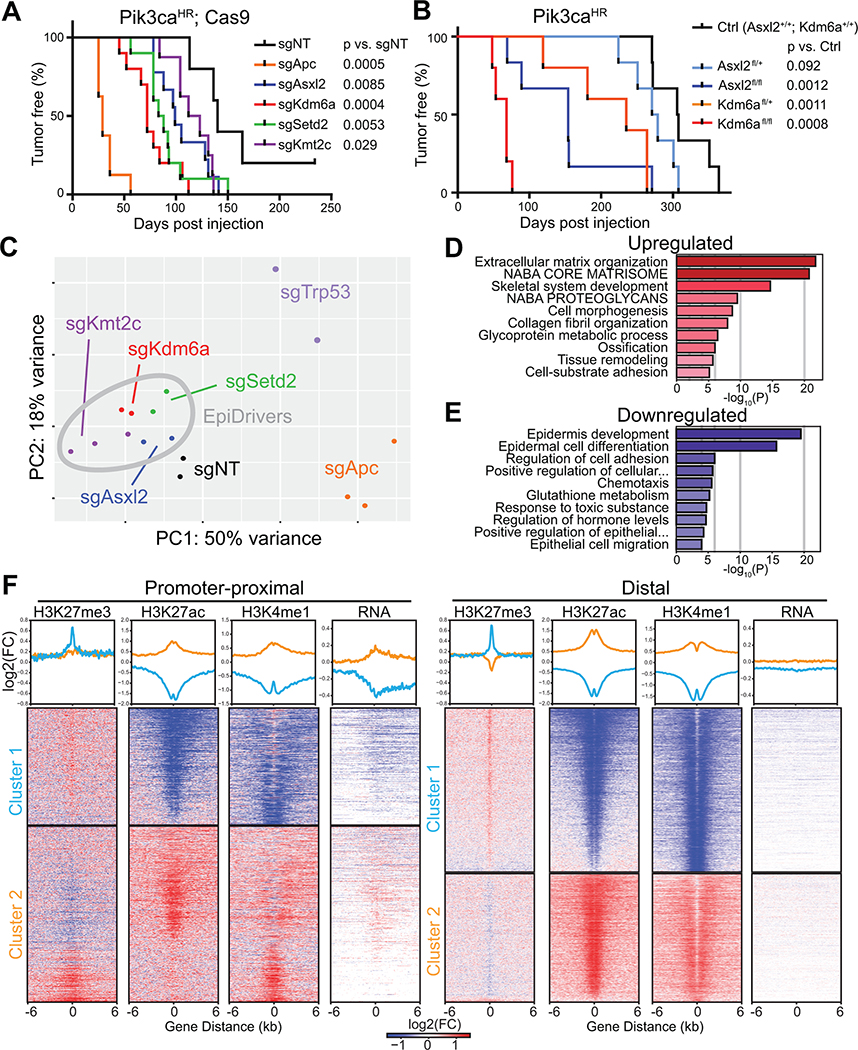
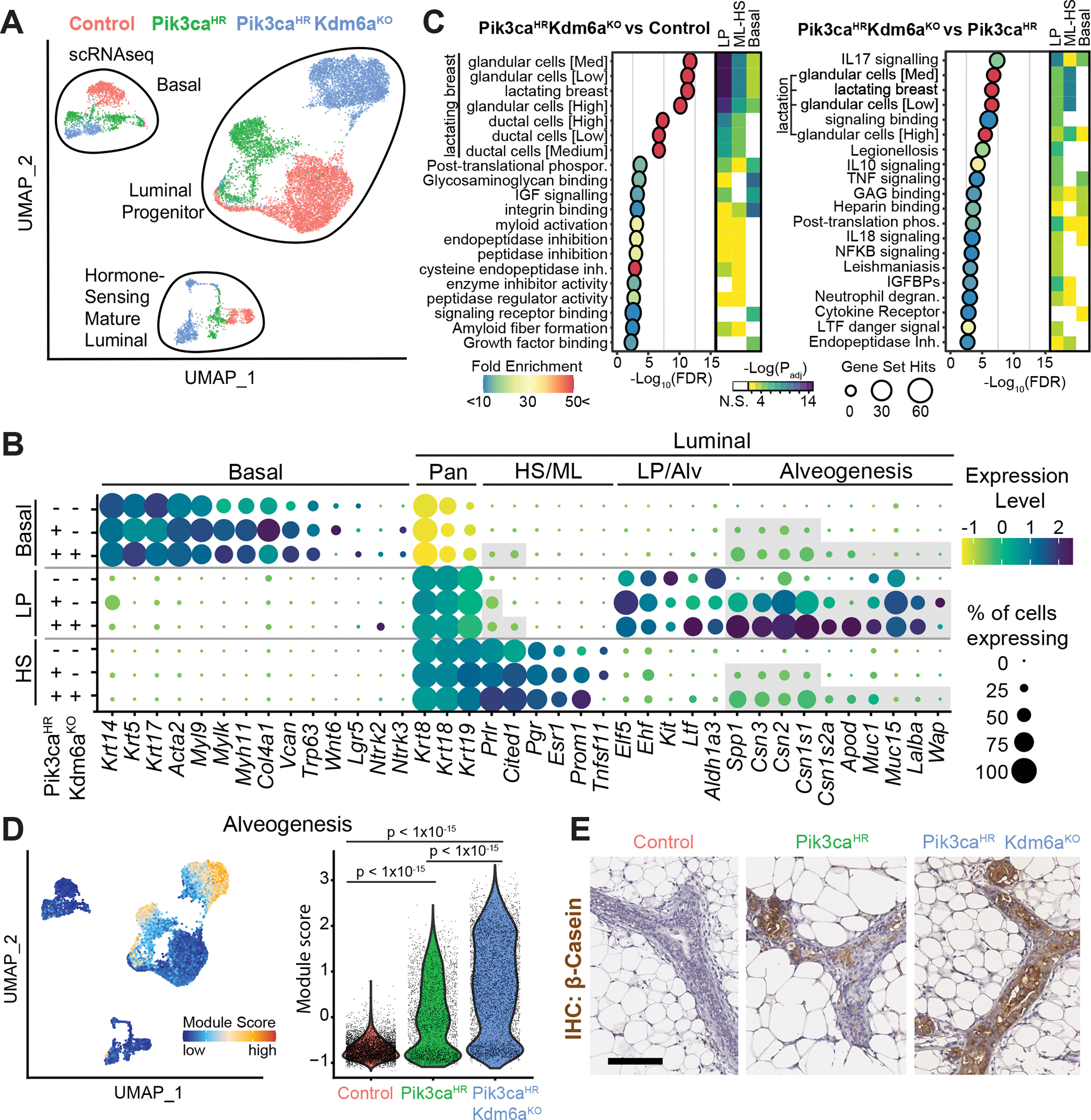
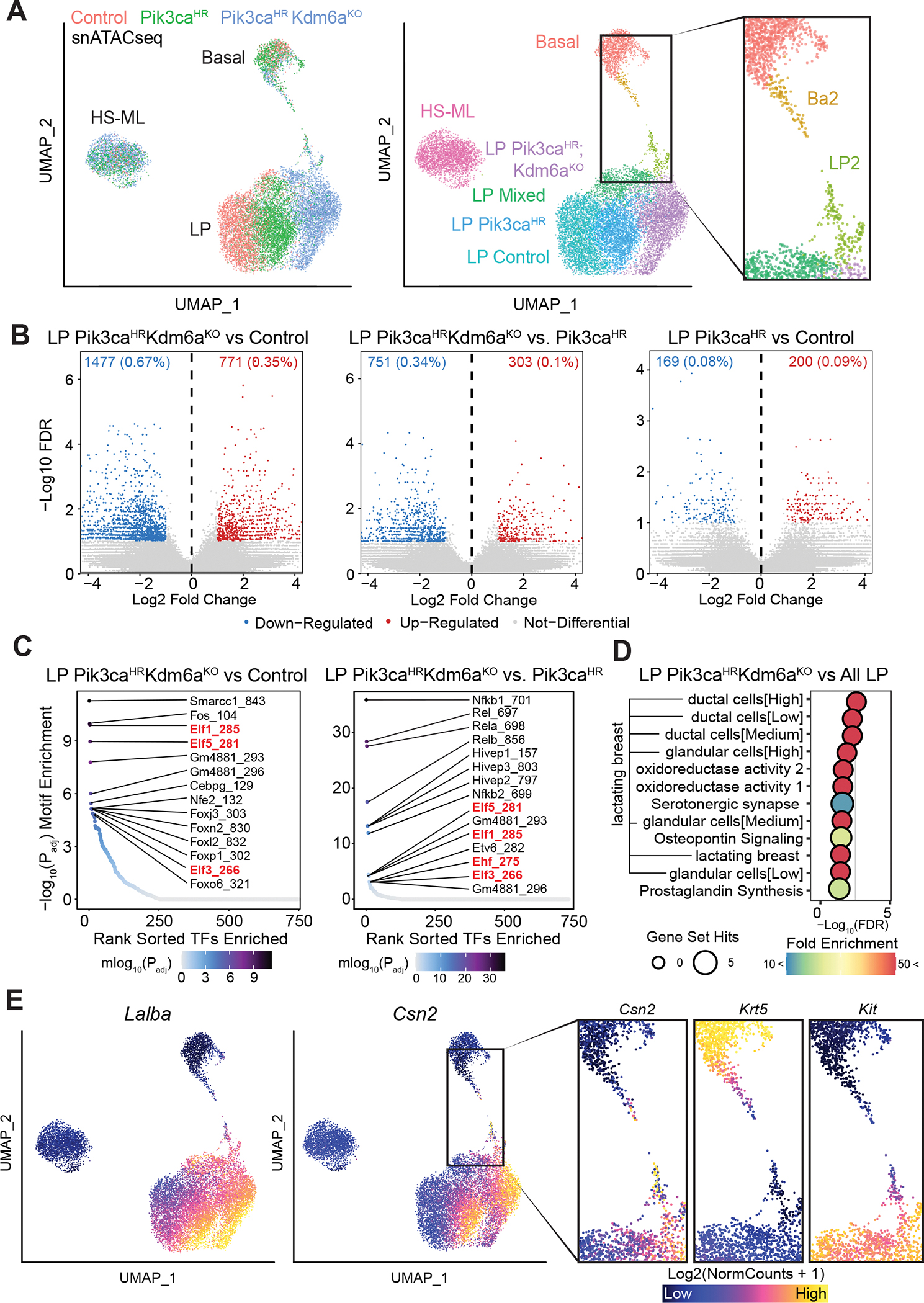
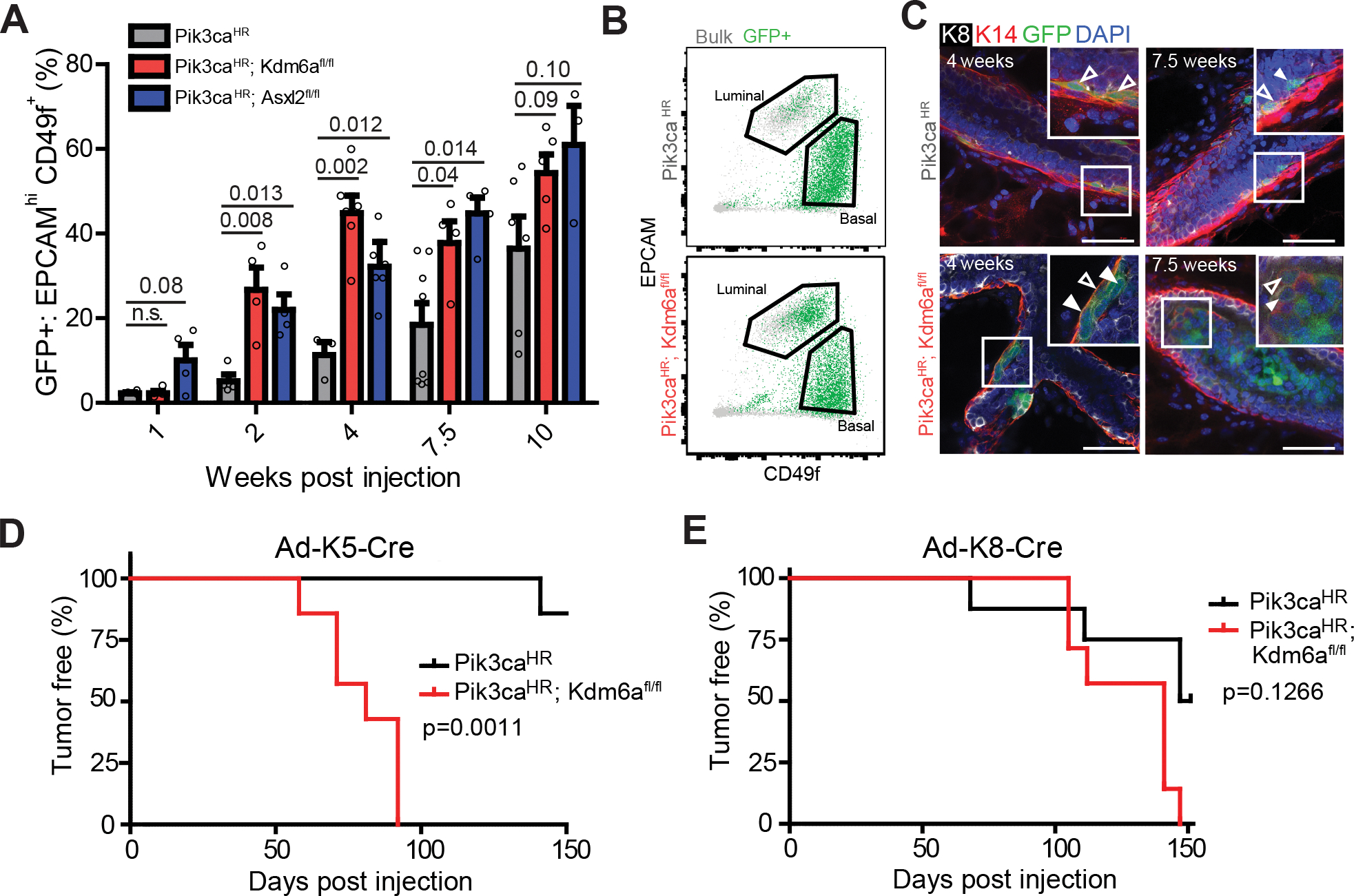
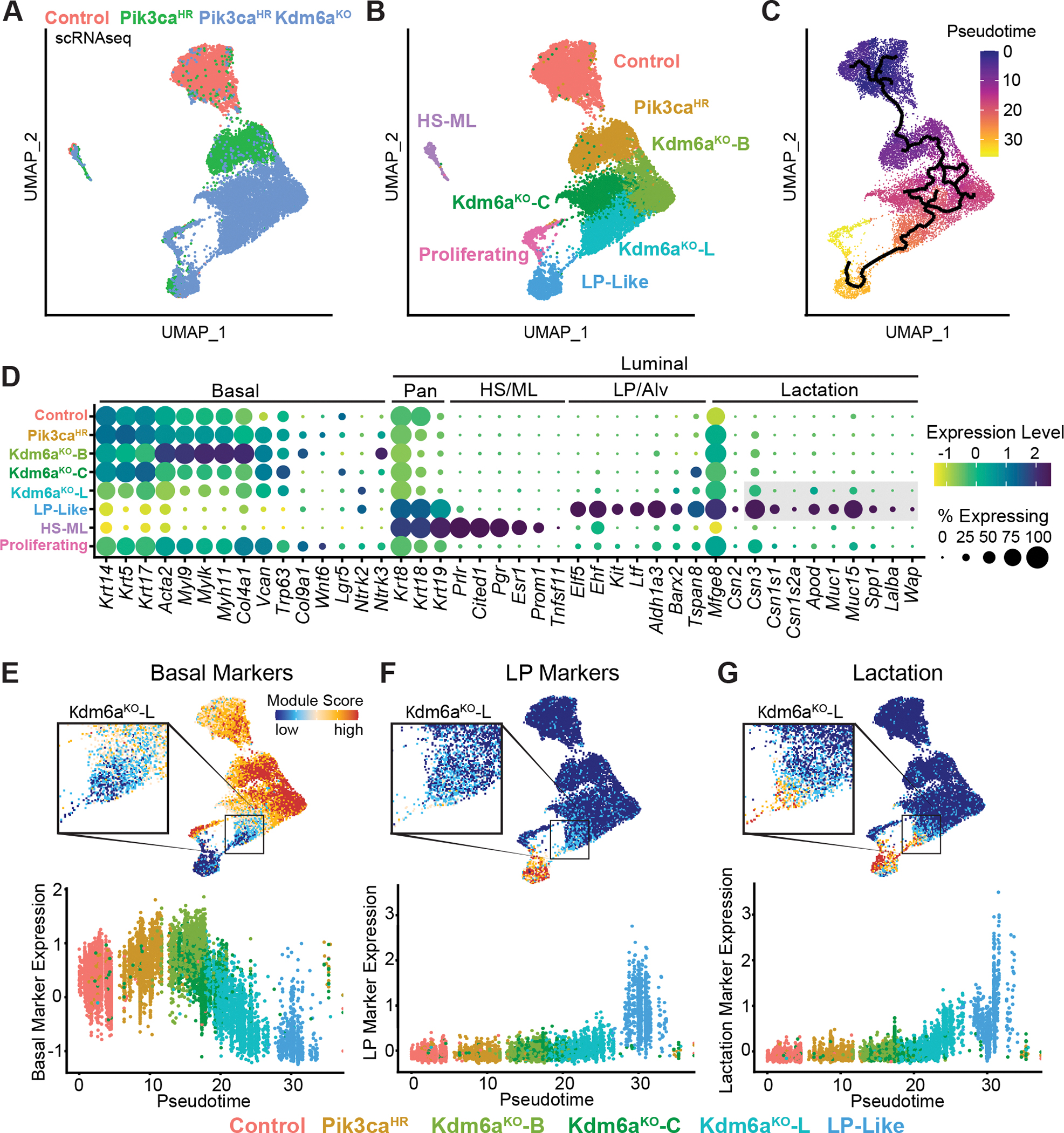
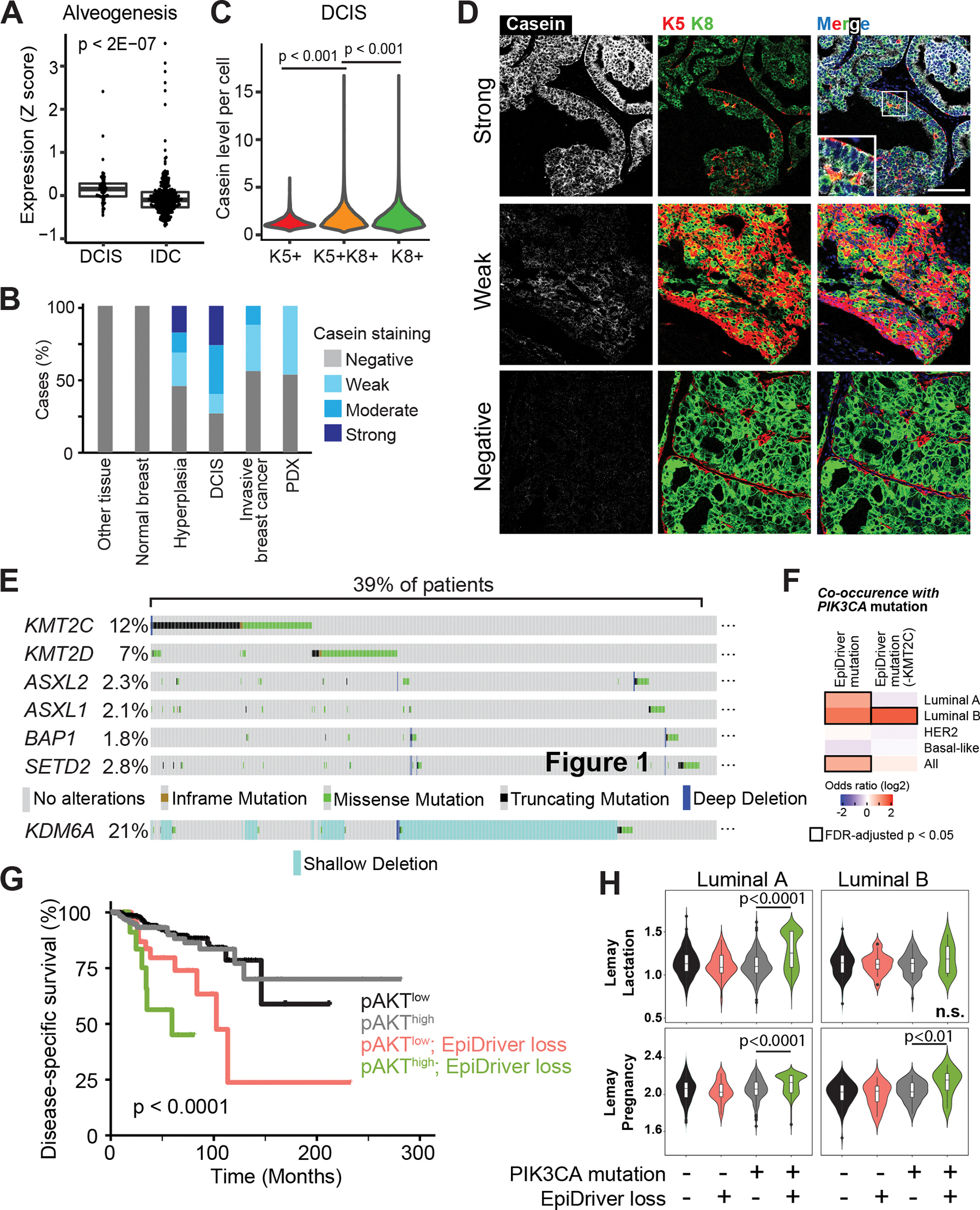
Systematically investigating the scores of genes mutated in cancer and discerning disease drivers from inconsequential bystanders is a prerequisite for precision medicine but remains challenging. Here, we developed a somatic CRISPR/Cas9 mutagenesis screen to study 215 recurrent "long-tail" breast cancer genes, which revealed epigenetic regulation as a major tumor-suppressive mechanism. We report that components of the BAP1 and COMPASS-like complexes, including KMT2C/D, KDM6A, BAP1, and ASXL1/2 ("EpiDrivers"), cooperate with PIK3CAH1047R to transform mouse and human breast epithelial cells. Mechanistically, we find that activation of PIK3CAH1047R and concomitant EpiDriver loss triggered an alveolar-like lineage conversion of basal mammary epithelial cells and accelerated formation of luminal-like tumors, suggesting a basal origin for luminal tumors. EpiDriver mutations are found in ∼39% of human breast cancers, and ∼50% of ductal carcinoma in situ express casein, suggesting that lineage infidelity and alveogenic mimicry may significantly contribute to early steps of breast cancer etiology.
Significance: Infrequently mutated genes comprise most of the mutational burden in breast tumors but are poorly understood. In vivo CRISPR screening identified functional tumor suppressors that converged on epigenetic regulation. Loss of epigenetic regulators accelerated tumorigenesis and revealed lineage infidelity and aberrant expression of alveogenesis genes as potential early events in tumorigenesis. This article is highlighted in the In This Issue feature, p. 2711.
©2022 American Association for Cancer Research.
Conflict of interest statement
The authors declare no potential conflicts of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
