

Single-cell atlas of human liver development reveals pathways directing hepatic cell fates
- PMID: 36109670
- PMCID: PMC7617064
- DOI: 10.1038/s41556-022-00989-7
Single-cell atlas of human liver development reveals pathways directing hepatic cell fates
Abstract
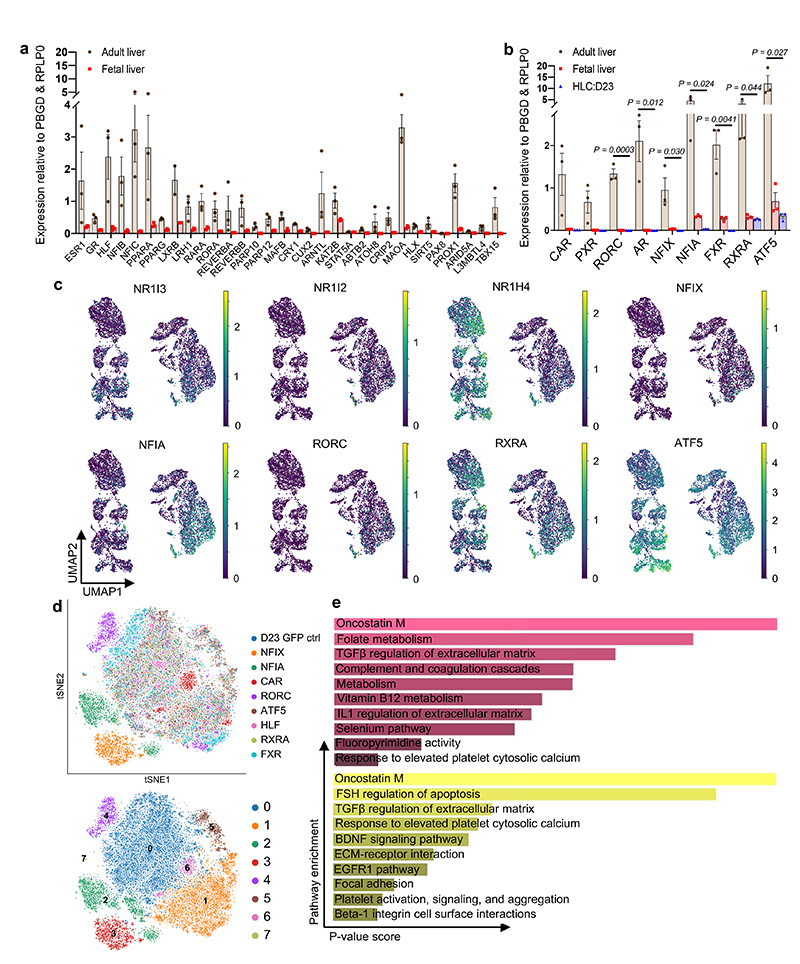
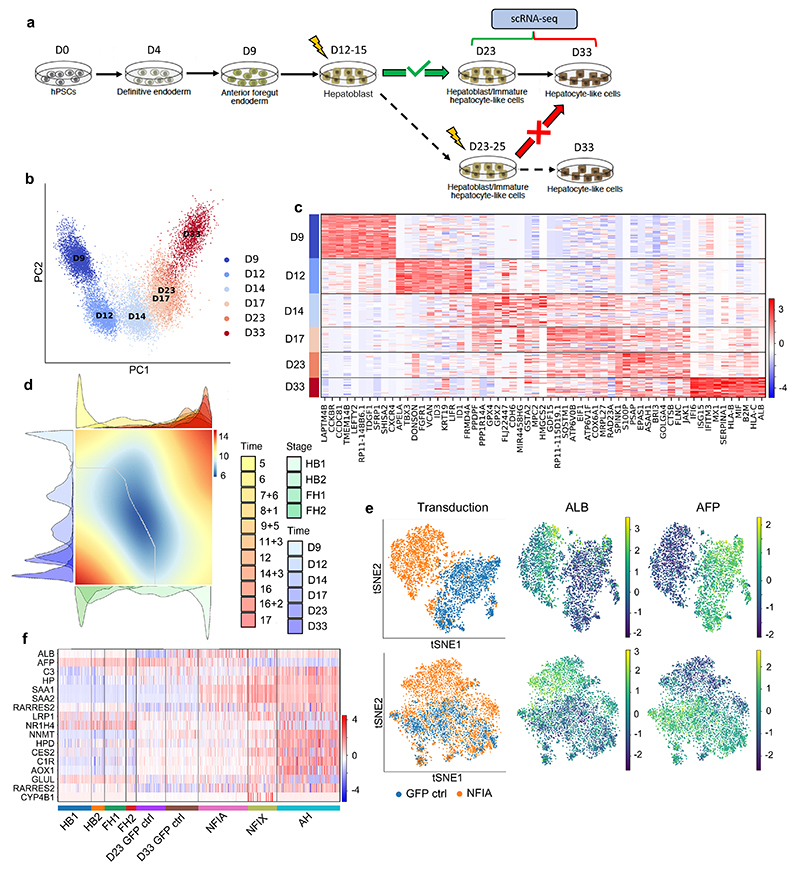
The liver has been studied extensively due to the broad number of diseases affecting its vital functions. However, therapeutic advances have been hampered by the lack of knowledge concerning human hepatic development. Here, we addressed this limitation by describing the developmental trajectories of different cell types that make up the human liver at single-cell resolution. These transcriptomic analyses revealed that sequential cell-to-cell interactions direct functional maturation of hepatocytes, with non-parenchymal cells playing essential roles during organogenesis. We utilized this information to derive bipotential hepatoblast organoids and then exploited this model system to validate the importance of signalling pathways in hepatocyte and cholangiocyte specification. Further insights into hepatic maturation also enabled the identification of stage-specific transcription factors to improve the functionality of hepatocyte-like cells generated from human pluripotent stem cells. Thus, our study establishes a platform to investigate the basic mechanisms directing human liver development and to produce cell types for clinical applications.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
All authors declare no competing interests.
Figures
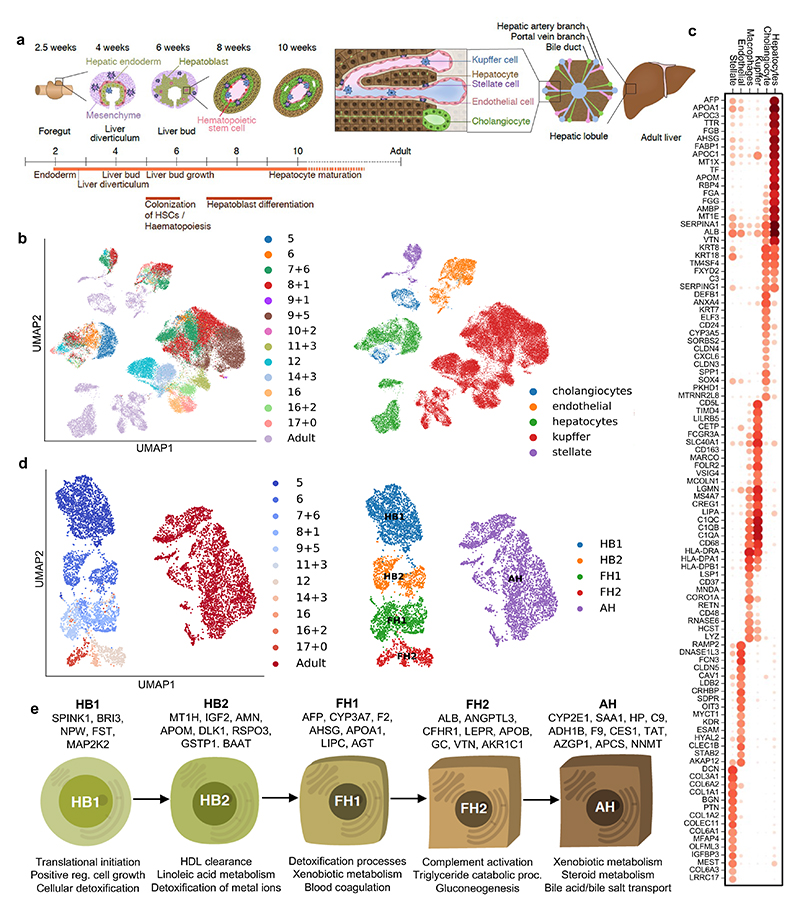
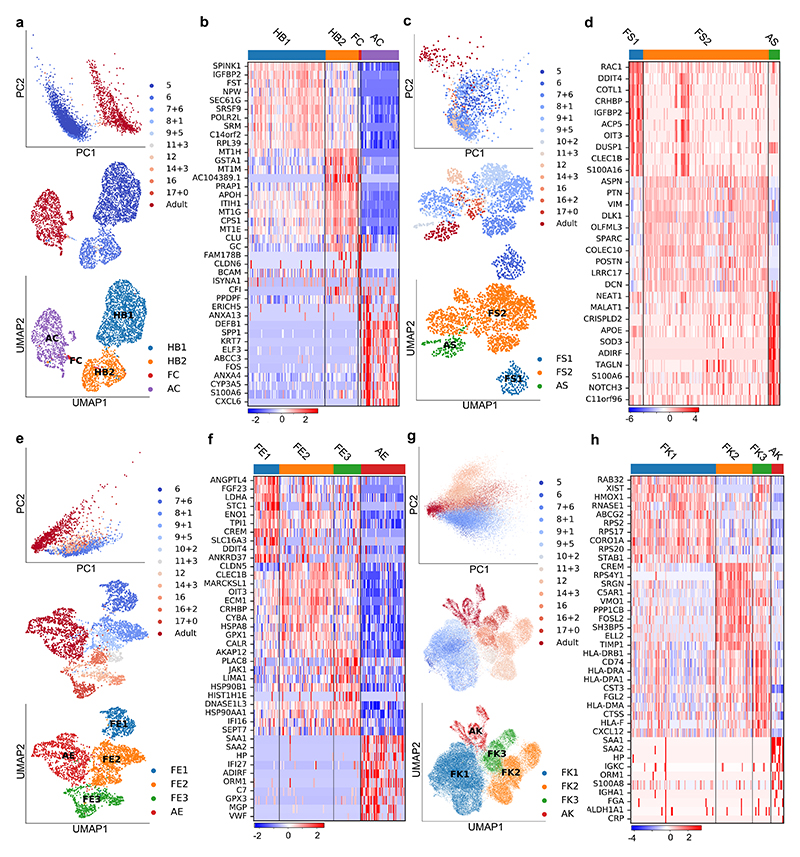
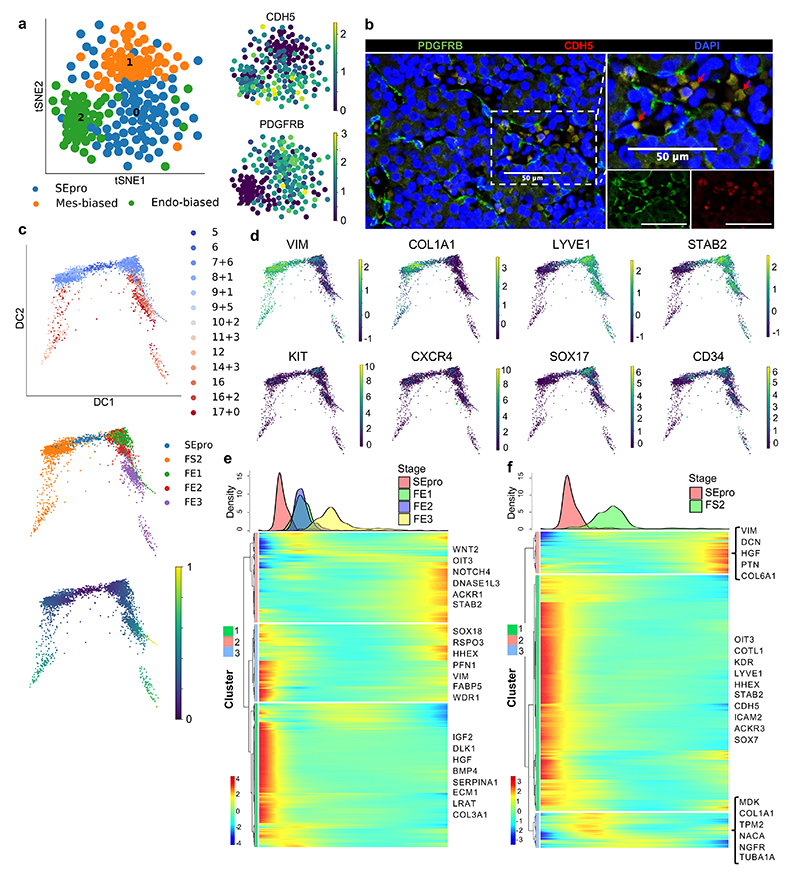
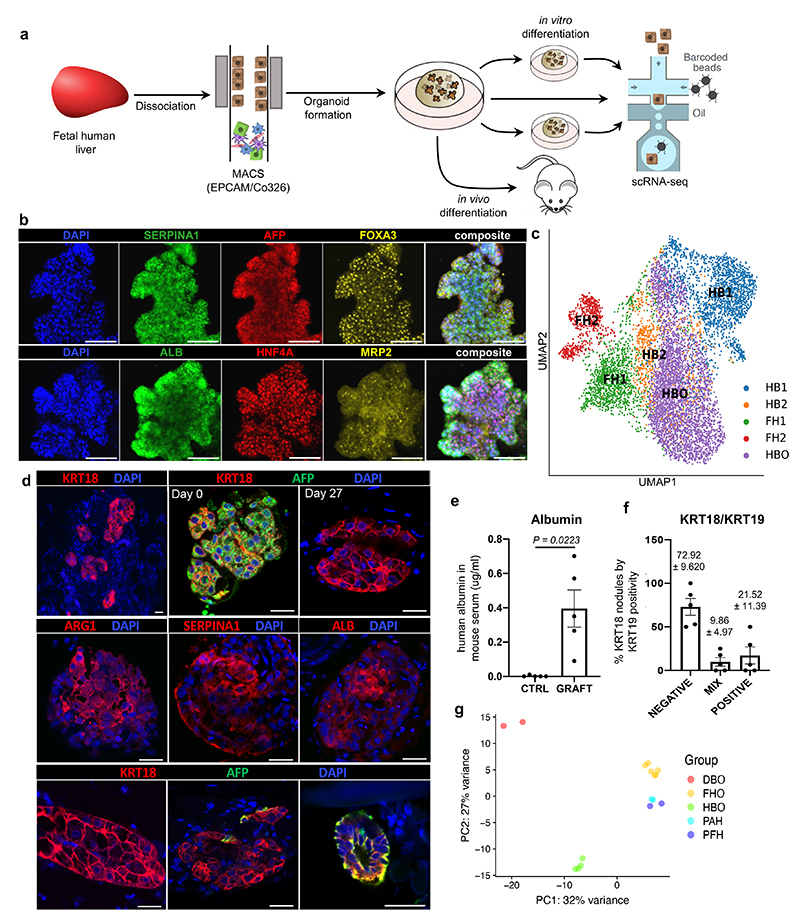
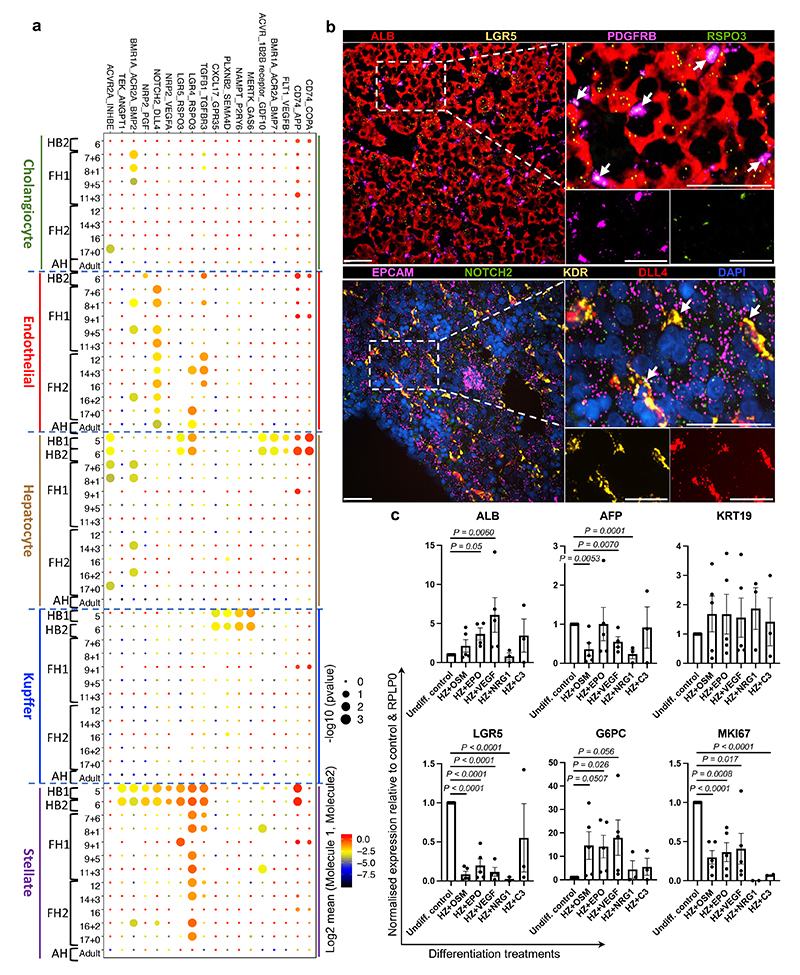
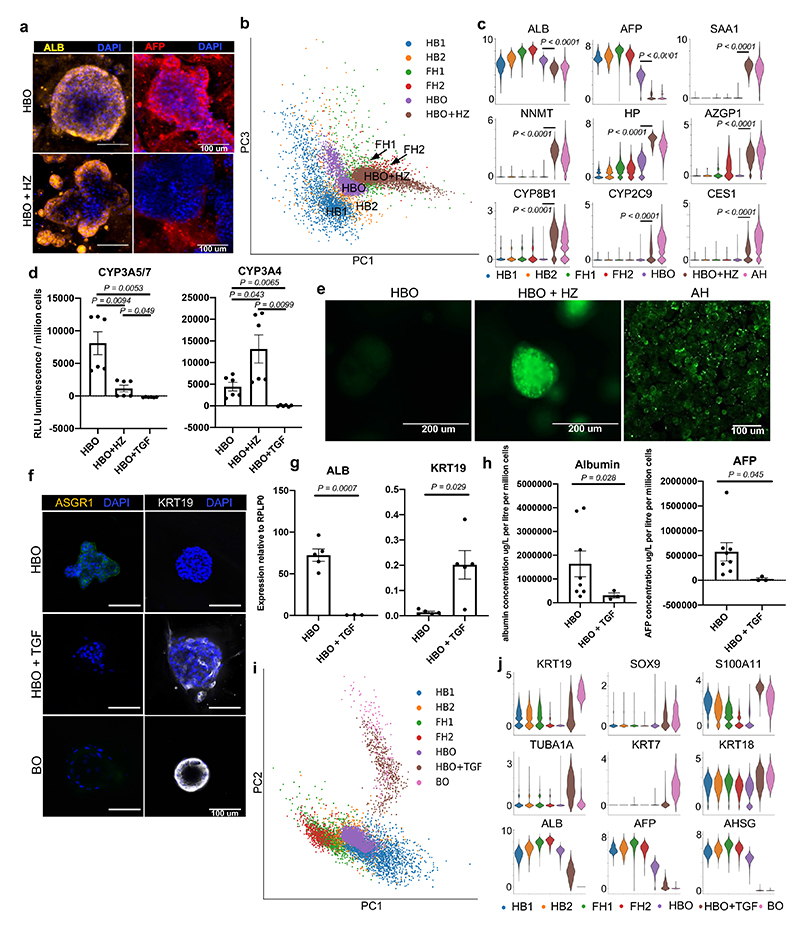
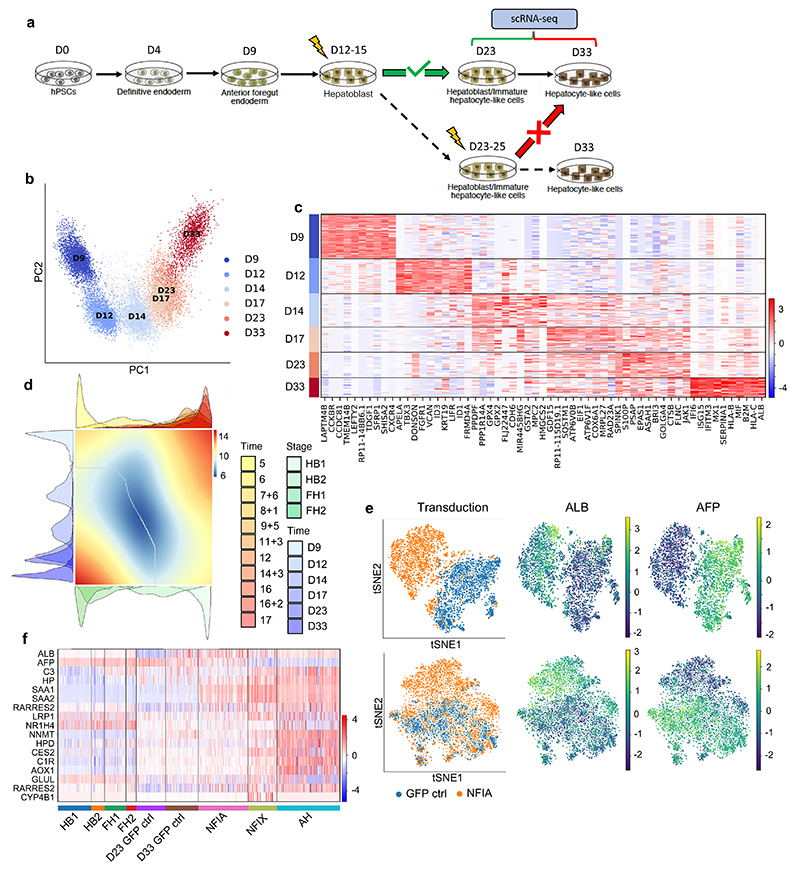
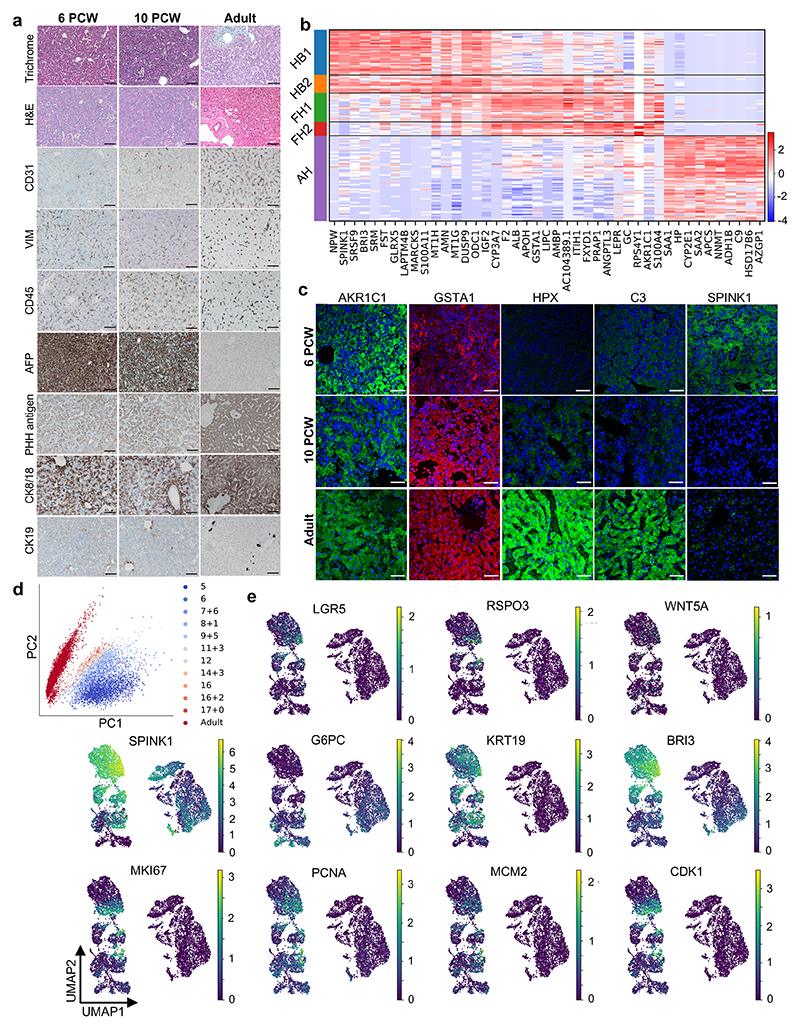
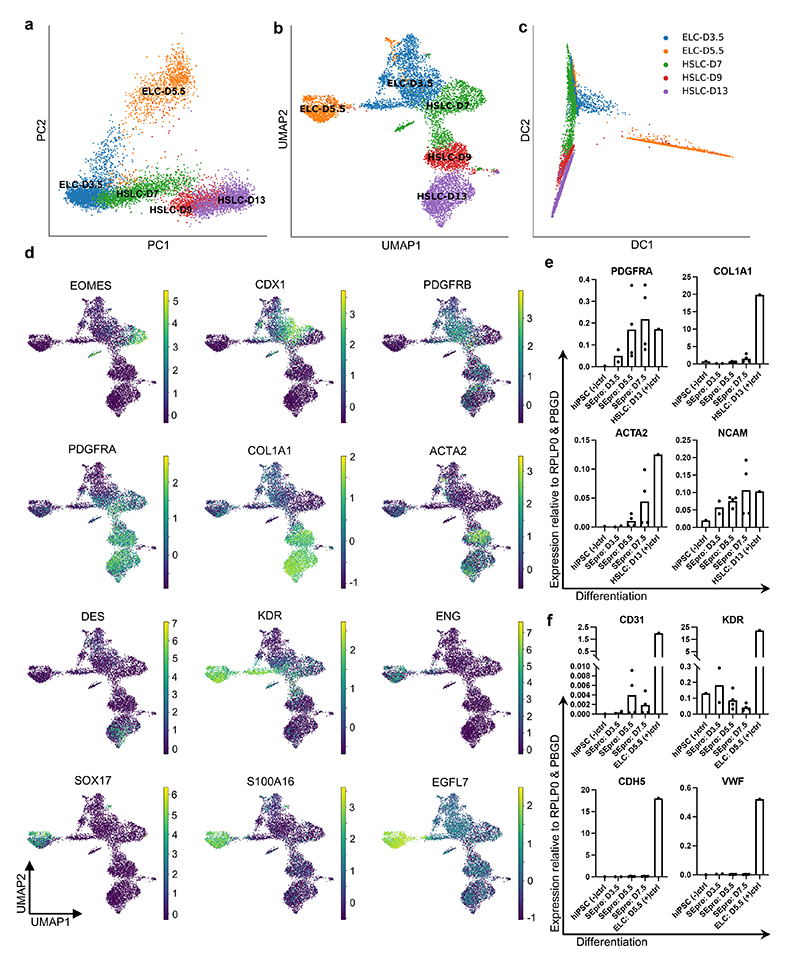
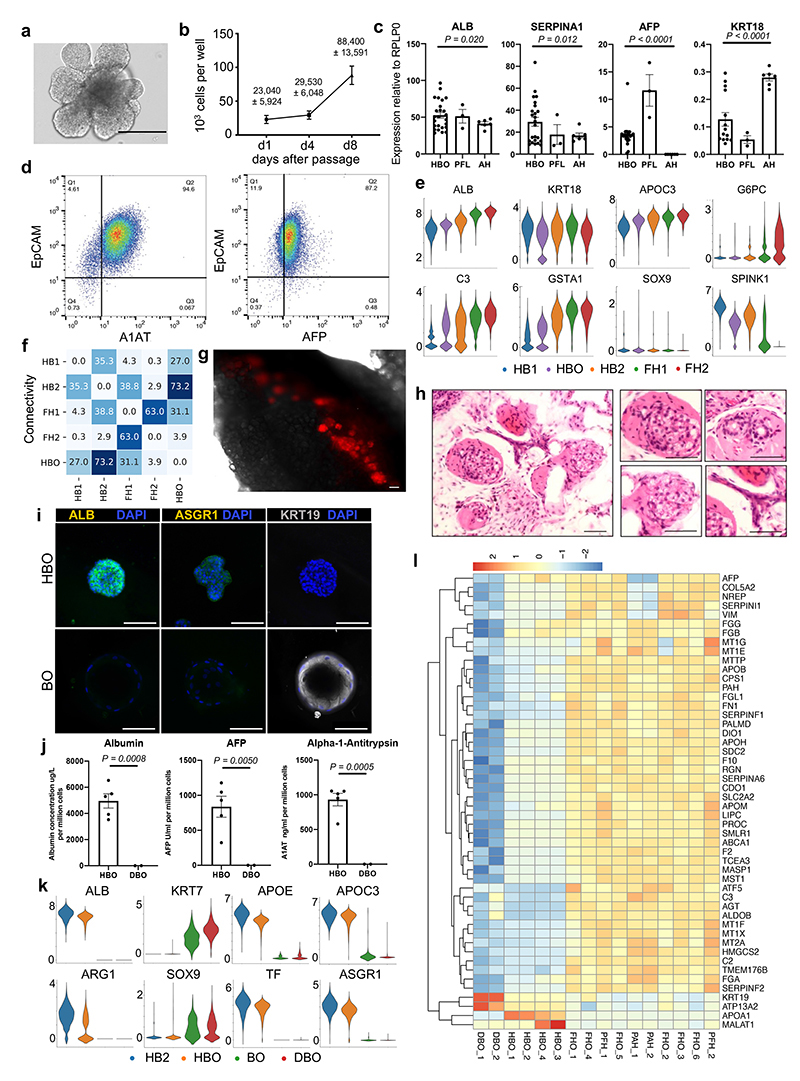
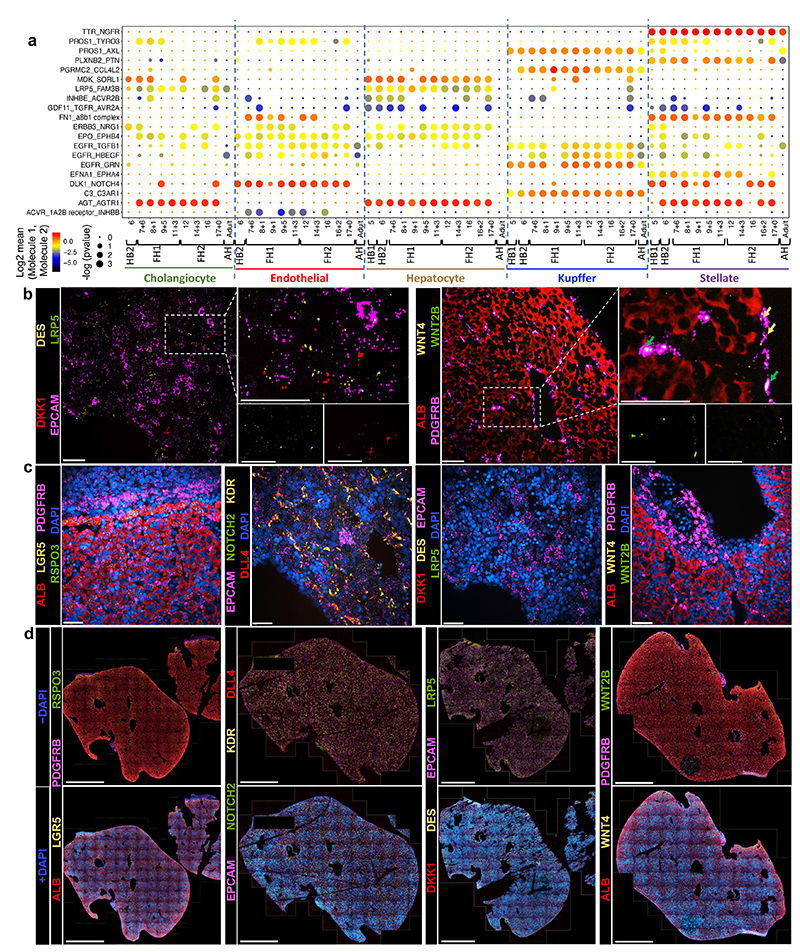
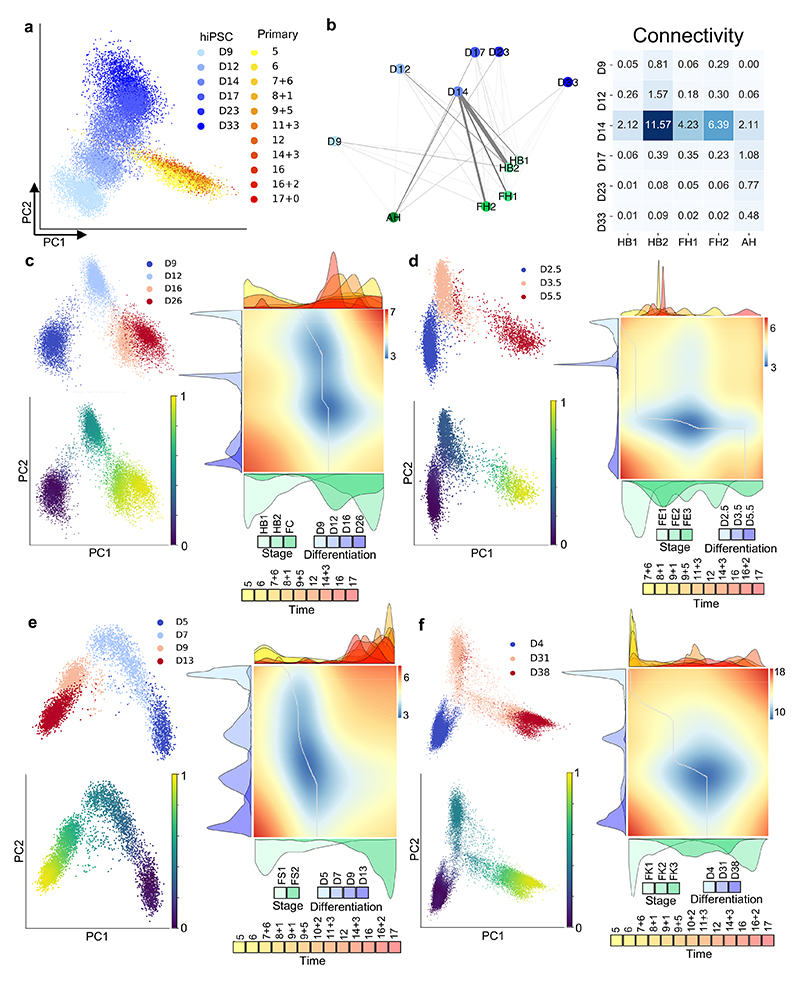
References
-
- Bilzer M, Roggel F, Gerbes AL. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006;26:1175–1186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- NC/N001540/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- DH_/Department of Health/United Kingdom
- 206194/WT_/Wellcome Trust/United Kingdom
- NC/R001987/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- NC/T2T0419/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
