

CDK12 regulates co-transcriptional splicing and RNA turnover in human cells
- PMID: 36111258
- PMCID: PMC9468413
- DOI: 10.1016/j.isci.2022.105030
CDK12 regulates co-transcriptional splicing and RNA turnover in human cells
Abstract
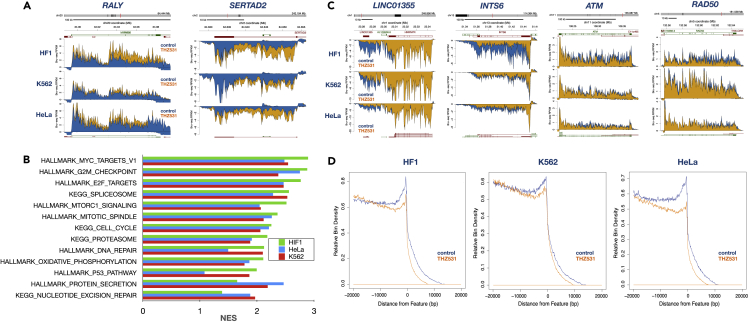
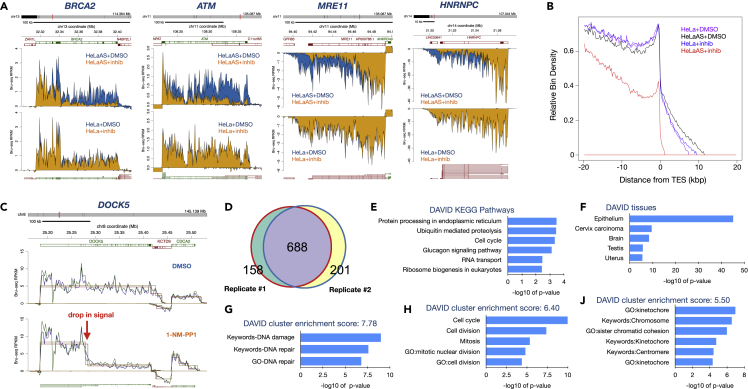
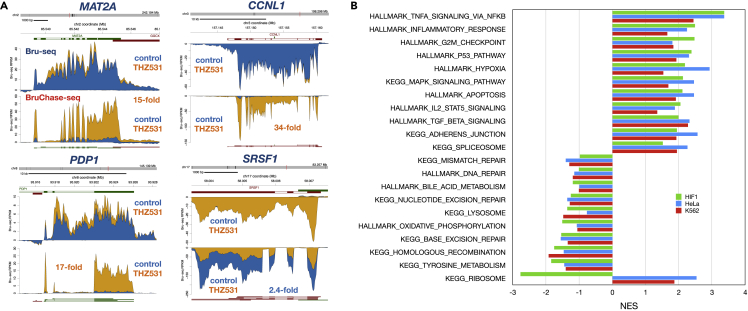
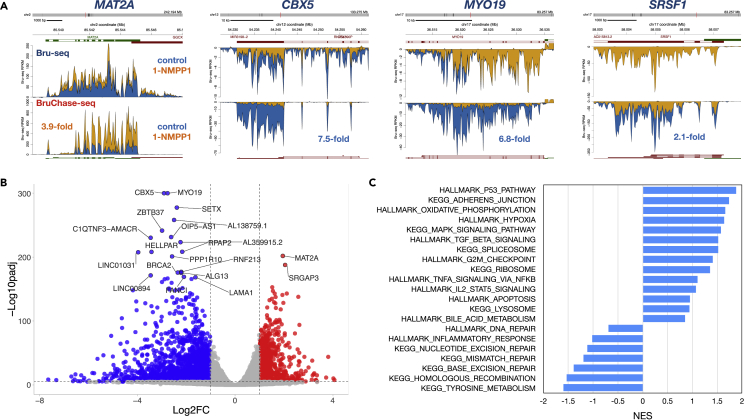
The cyclin-dependent kinase CDK12 has garnered interest as a cancer therapeutic target as DNA damage response genes are particularly suppressed by loss of CDK12 activity. In this study, we assessed the acute effects of CDK12 inhibition on transcription and RNA processing using nascent RNA Bru-seq and BruChase-seq. Acute transcriptional changes were overall small after CDK12 inhibition but over 600 genes showed intragenic premature termination, including DNA repair and cell cycle genes. Furthermore, many genes showed reduced transcriptional readthrough past the end of genes in the absence of CDK12 activity. RNA turnover was dramatically affected by CDK12 inhibition and importantly, caused increased degradation of many transcripts from DNA damage response genes. We also show that co-transcriptional splicing was suppressed by CDK12 inhibition. Taken together, these studies reveal the roles of CDK12 in regulating transcription elongation, transcription termination, co-transcriptional splicing, and RNA turnover.
Keywords: Biological sciences; cell biology; molecular biology.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interest.
Figures



References
-
- Ameur A., Zaghlool A., Halvardson J., Wetterbom A., Gyllensten U., Cavelier L., Feuk L. Total RNA sequencing reveals nascent transcription and widespread co-transcriptional splicing in the human brain. Nat. Struct. Mol. Biol. 2011;18:1435–1440. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
