

Activation of TrkB-Akt signaling rescues deficits in a mouse model of SCA6
- PMID: 36112675
- PMCID: PMC9481119
- DOI: 10.1126/sciadv.abh3260
Activation of TrkB-Akt signaling rescues deficits in a mouse model of SCA6
Abstract
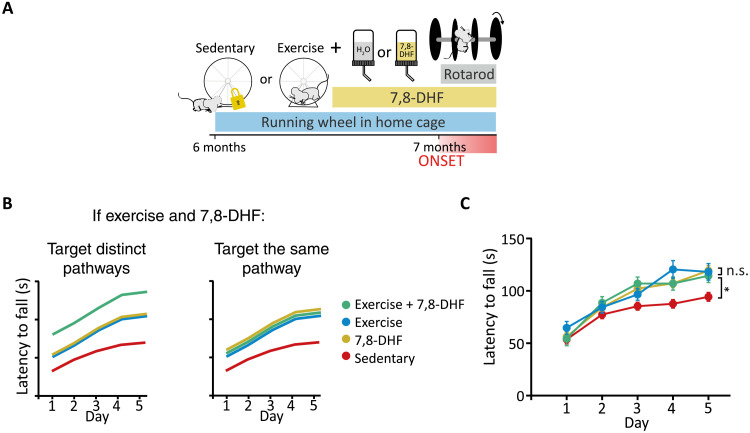
Spinocerebellar ataxia type 6 (SCA6) is a neurodegenerative disease resulting in motor coordination deficits and cerebellar pathology. Expression of brain-derived neurotrophic factor (BDNF) is reduced in postmortem tissue from SCA6 patients. Here, we show that levels of cerebellar BDNF and its receptor, tropomyosin receptor kinase B (TrkB), are reduced at an early disease stage in a mouse model of SCA6 (SCA684Q/84Q). One month of exercise elevated cerebellar BDNF expression and improved ataxia and cerebellar Purkinje cell firing rate deficits. A TrkB agonist, 7,8-dihydroxyflavone (7,8-DHF), likewise improved motor coordination and Purkinje cell firing rate and elevated downstream Akt signaling. Prolonged 7,8-DHF administration persistently improved ataxia when treatment commenced near disease onset but was ineffective when treatment was started late. These data suggest that 7,8-DHF, which is orally bioavailable and crosses the blood-brain barrier, is a promising therapeutic for SCA6 and argue for the importance of early intervention for SCA6.
Figures







References
-
- Zhuchenko O., Bailey J., Bonnen P., Ashizawa T., Stockton D. W., Amos C., Dobyns W. B., Subramony S. H., Zoghbi H. Y., Lee C. C., Autosomal dominant cerebellar ataxia (SCA6) associated with small polyglutamine expansions in the alpha 1A-voltage-dependent calcium channel. Nat. Genet. 15, 62–69 (1997). - PubMed
-
- Solodkin A., Gomez C. M., Spinocerebellar ataxia type 6. Handb. Clin. Neurol. 103, 461–473 (2012). - PubMed
-
- Watase K., Barrett C. F., Miyazaki T., Ishiguro T., Ishikawa K., Hu Y., Unno T., Sun Y., Kasai S., Watanabe M., Gomez C. M., Mizusawa H., Tsien R. W., Zoghbi H. Y., Spinocerebellar ataxia type 6 knockin mice develop a progressive neuronal dysfunction with age-dependent accumulation of mutant CaV2.1 channels. Proc. Natl. Acad. Sci. U.S.A. 105, 11987–11992 (2008). - PMC - PubMed
-
- Zuccato C., Cattaneo E., Brain-derived neurotrophic factor in neurodegenerative diseases. Nat. Rev. Neurol. 5, 311–322 (2009). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
