

The effects of β-catenin on cardiomyogenesis via Islet-1 and MLIP ubiquitination
- PMID: 36112854
- PMCID: PMC9742745
- DOI: 10.1177/15353702221119792
The effects of β-catenin on cardiomyogenesis via Islet-1 and MLIP ubiquitination
Abstract
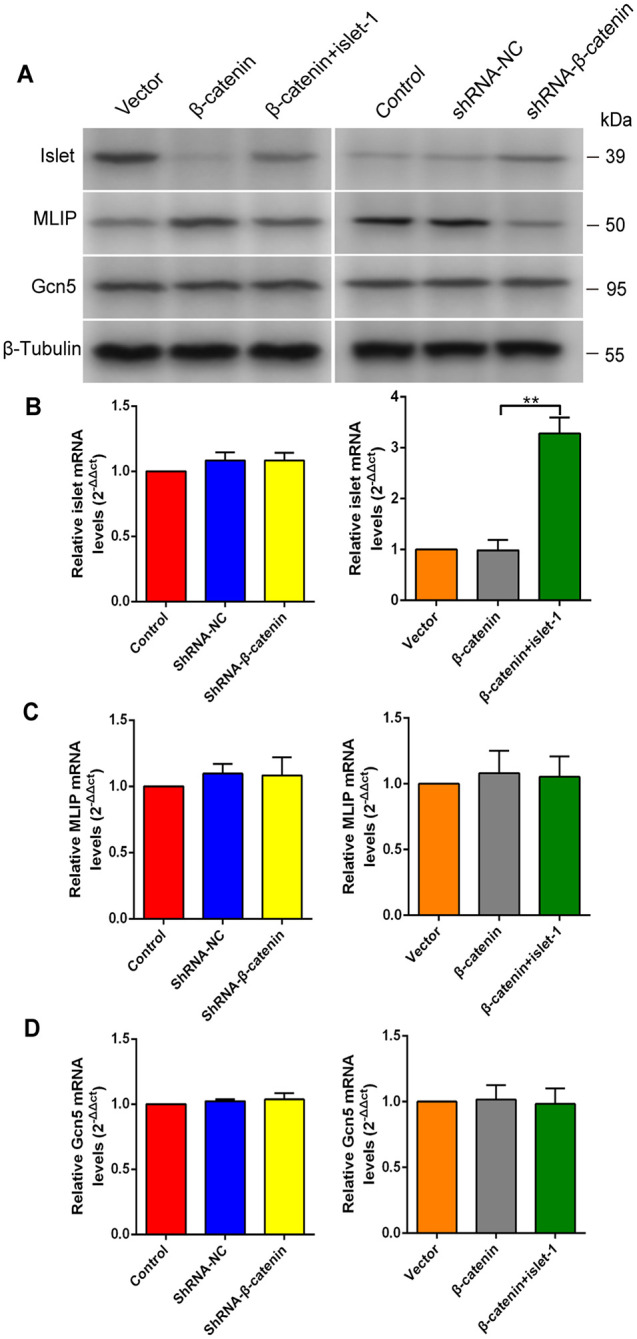
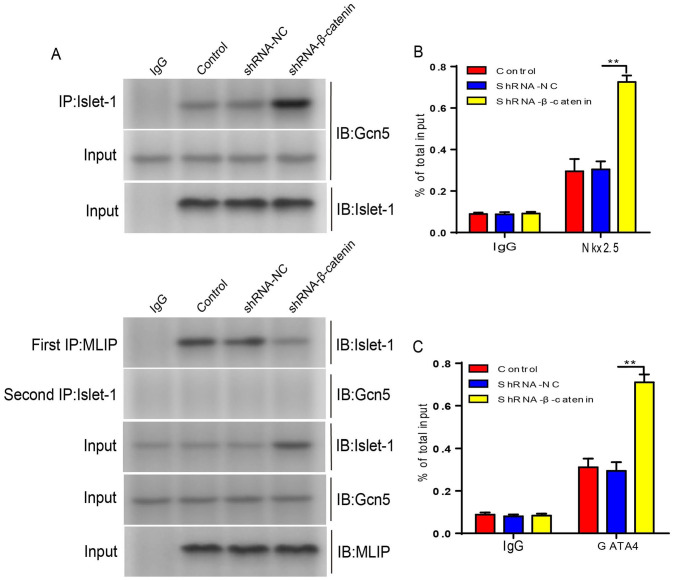
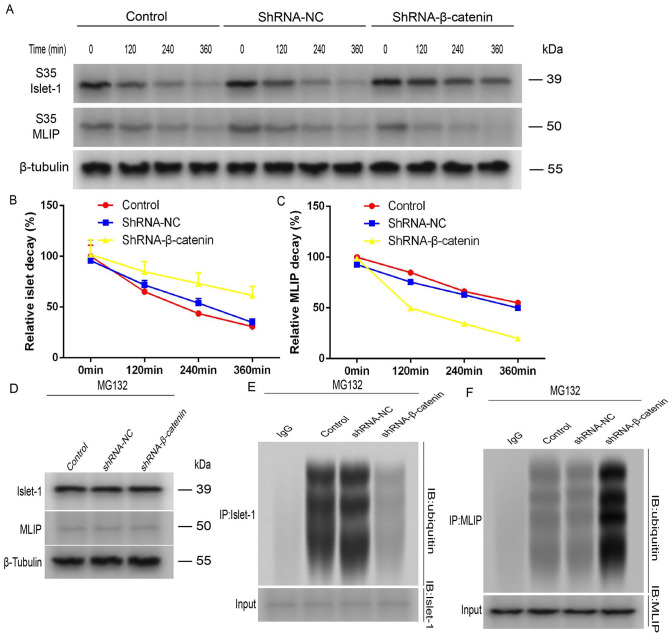
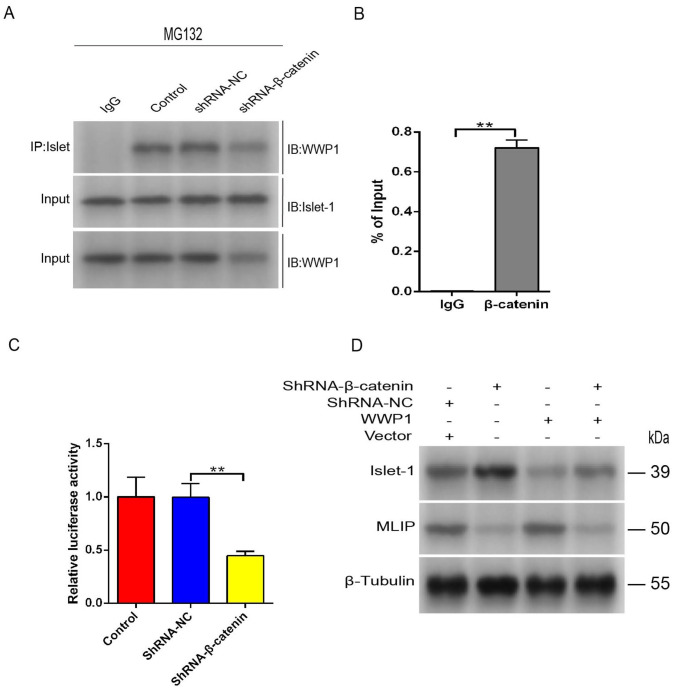
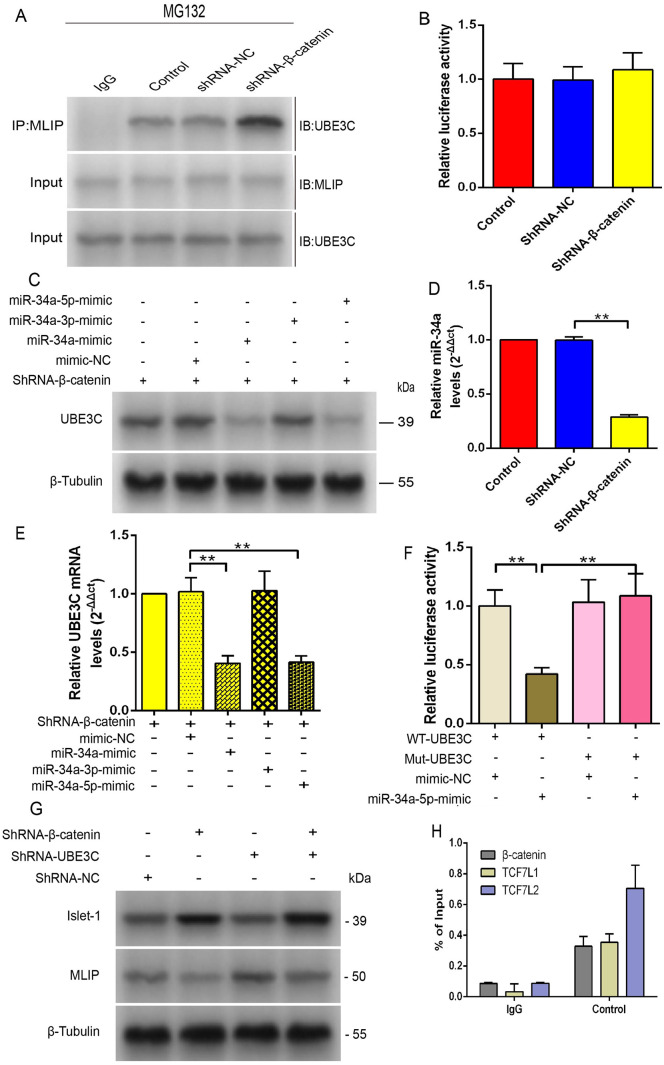
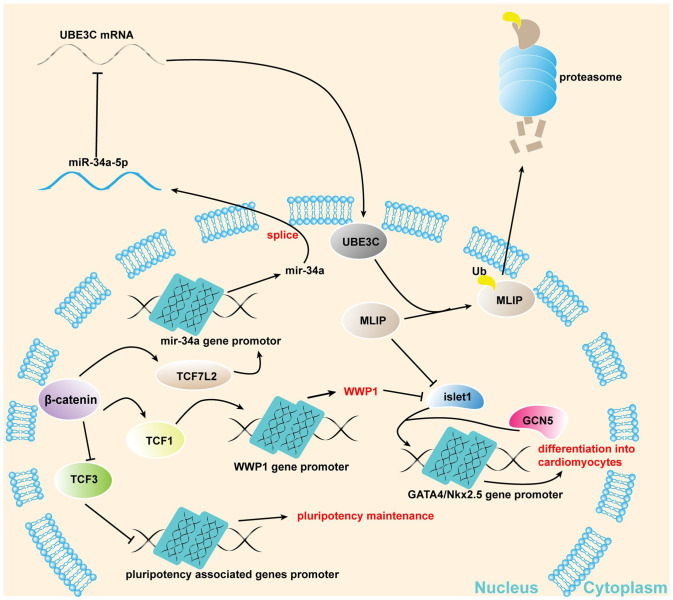
Mesenchymal stem cells (MSCs) can treat myocardial injury-related diseases by differentiating into cardiomyocytes. Islet-1 plays an essential role in cardiac maturation. We have discovered that Islet-1 plays a crucial role in the histone acetylation regulation in this process. In addition, to increase GATA4/Nkx2.5 expression, Islet-1 may bind to Gcn5 and then guide Gcn5 to the GATA4/Nkx2.5 promoters, thereby facilitating the differentiation of MSCs into cardiomyocytes. Islet-1 is an important factor in the maturation of the heart. We have previously found that the pivotal factor in histone acetylation regulation in this process is Islet-1. Furthermore, Islet-1 and Gcn5 may boost GATA4/Nkx2.5 expression, which in turn promotes cardiomyocyte differentiation from MSCs. But the molecular mechanism of Islet-1 binding to GCN5 has not been elucidated. In this study, we found that the competitive binding relationship between Islet-1 and MLIP and GCN5 affected myocardial differentiation. The key enzymes of ubiquitination modification of MLIP and Islet-1 are UBE3C and WWP1, respectively. When short hairpin RNA (shRNA) was used to inhibit β-catenin expression, we found that the expression of UBE3C was upregulated, modifying MLIP ubiquitination and reducing its expression, and it upregulated Islet-1 by inhibiting the expression of WWP1. By using the chromatin immunoprecipitation (ChIP) and luciferase reporter system, we found that when MLIP binds to Islet-1, it significantly inhibits the transcriptional activity of Islet-1. In summary, our results show that decreasing β-catenin regulates the ubiquitination of Islet-1 and MLIP, affecting their expression, reducing the amount of Islet-1 binding to MLIP, and increasing the amount of binding to GCN5 in the nucleus. Therefore, the transcriptional activity of Islet-1 is significantly activated, inducing C3H10T1/2 cells to differentiate into myocytes. Further knowledge of biochemical pathways, including molecular signaling pathways, can provide more insights into the myocardial differentiation mechanism of MSCs.
Keywords: C3H10T1/2 cells; Islet-1; MLIP; cardiomyogenesis; ubiquitination; β-catenin.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
Similar articles
-
Islet-1 synergizes with Gcn5 to promote MSC differentiation into cardiomyocytes.Sci Rep. 2020 Feb 4;10(1):1817. doi: 10.1038/s41598-020-58387-8. Sci Rep. 2020. PMID: 32019948 Free PMC article.
-
A role for Gcn5 in cardiomyocyte differentiation of rat mesenchymal stem cells.Mol Cell Biochem. 2010 Dec;345(1-2):309-16. doi: 10.1007/s11010-010-0586-3. Epub 2010 Sep 12. Mol Cell Biochem. 2010. PMID: 20835911
-
Islet-1 induces the differentiation of mesenchymal stem cells into cardiomyocyte-like cells through the regulation of Gcn5 and DNMT-1.Mol Med Rep. 2017 May;15(5):2511-2520. doi: 10.3892/mmr.2017.6343. Epub 2017 Mar 16. Mol Med Rep. 2017. PMID: 28447752 Free PMC article.
-
Islet-1 promotes the cardiac-specific differentiation of mesenchymal stem cells through the regulation of histone acetylation.Int J Mol Med. 2014 May;33(5):1075-82. doi: 10.3892/ijmm.2014.1687. Epub 2014 Mar 6. Int J Mol Med. 2014. PMID: 24604334 Free PMC article.
-
WNT signaling promotes Nkx2.5 expression and early cardiomyogenesis via downregulation of Hdac1.Biochim Biophys Acta. 2009 Feb;1793(2):300-11. doi: 10.1016/j.bbamcr.2008.08.013. Epub 2008 Sep 11. Biochim Biophys Acta. 2009. PMID: 18851995
Cited by
-
Nkx2.5: a crucial regulator of cardiac development, regeneration and diseases.Front Cardiovasc Med. 2023 Dec 6;10:1270951. doi: 10.3389/fcvm.2023.1270951. eCollection 2023. Front Cardiovasc Med. 2023. PMID: 38124890 Free PMC article. Review.
References
-
- Tam PP, Parameswaran M, Kinder SJ, Weinberger RP. The allocation of epiblast cells to the embryonic heart and other mesodermal lineages: the role of ingression and tissue movement during gastrulation. Development 1997;124:1631–42 - PubMed
-
- Bu L, Jiang X, Martin-Puig S, Caron L, Zhu S, Shao Y, Roberts DJ, Huang PL, Domian IJ, Chien KR. Human ISL1 heart progenitors generate diverse multipotent cardiovascular cell lineages. Nature 2009;460:113–7 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases