

Structures reveal a key mechanism of WAVE regulatory complex activation by Rac1 GTPase
- PMID: 36114192
- PMCID: PMC9481577
- DOI: 10.1038/s41467-022-33174-3
Structures reveal a key mechanism of WAVE regulatory complex activation by Rac1 GTPase
Abstract
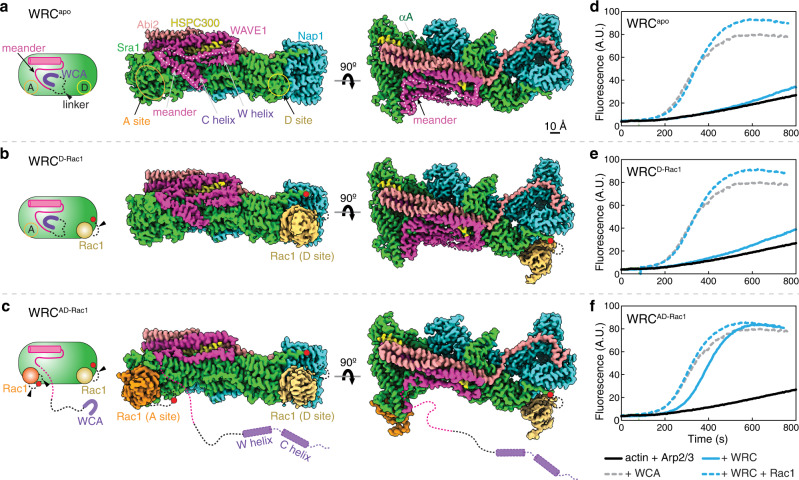
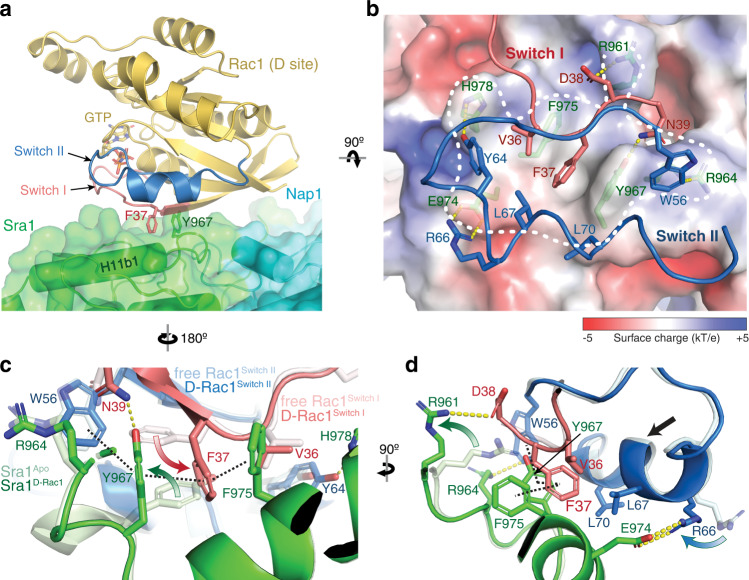
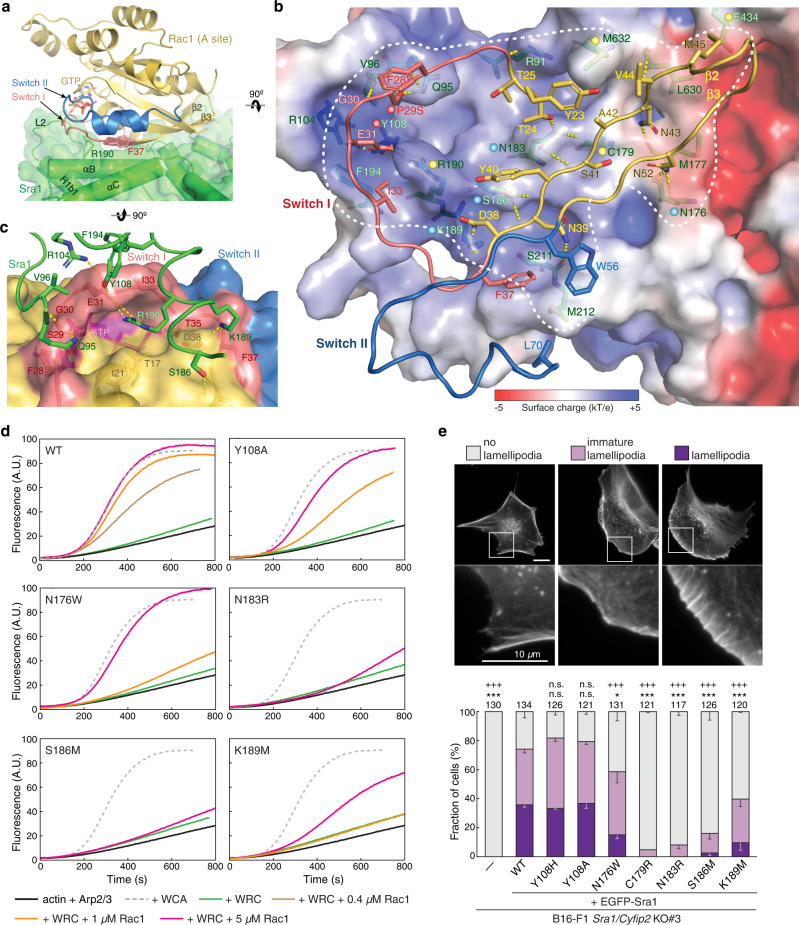
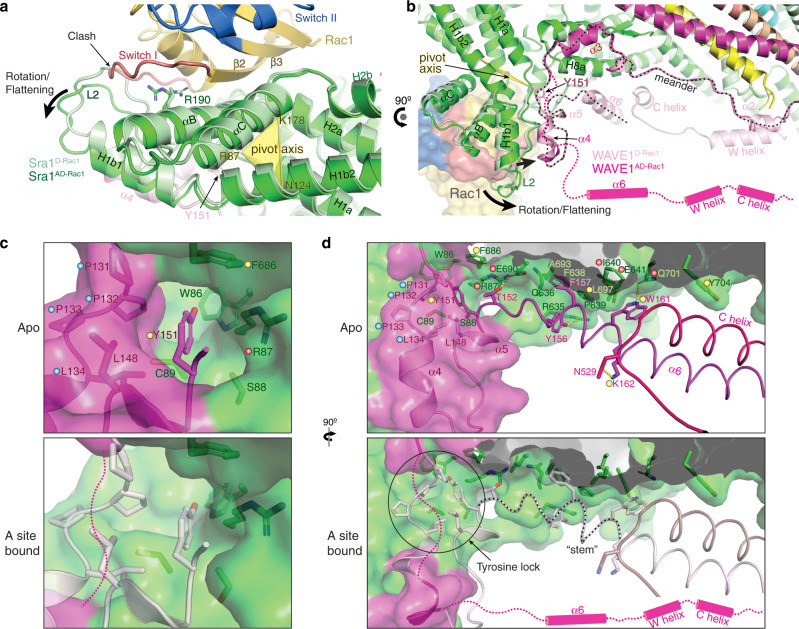
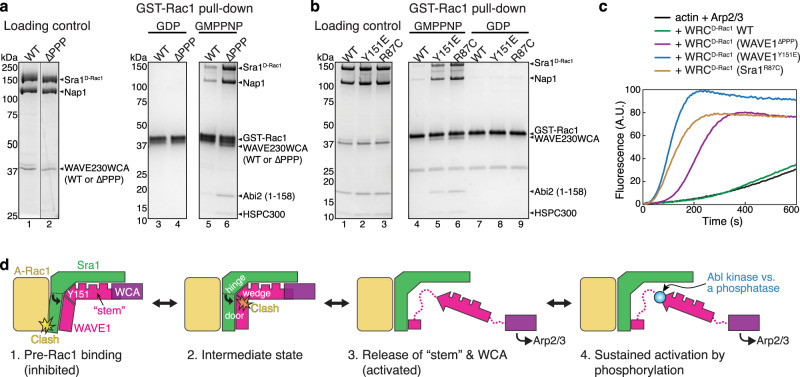
The Rho-family GTPase Rac1 activates the WAVE regulatory complex (WRC) to drive Arp2/3 complex-mediated actin polymerization in many essential processes. Rac1 binds to WRC at two distinct sites-the A and D sites. Precisely how Rac1 binds and how the binding triggers WRC activation remain unknown. Here we report WRC structures by itself, and when bound to single or double Rac1 molecules, at ~3 Å resolutions by cryogenic-electron microscopy. The structures reveal that Rac1 binds to the two sites by distinct mechanisms, and binding to the A site, but not the D site, drives WRC activation. Activation involves a series of unique conformational changes leading to the release of sequestered WCA (WH2-central-acidic) polypeptide, which stimulates the Arp2/3 complex to polymerize actin. Together with biochemical and cellular analyses, the structures provide a novel mechanistic understanding of how the Rac1-WRC-Arp2/3-actin signaling axis is regulated in diverse biological processes and diseases.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Rac1 GTPase activates the WAVE regulatory complex through two distinct binding sites.Elife. 2017 Sep 26;6:e29795. doi: 10.7554/eLife.29795. Elife. 2017. PMID: 28949297 Free PMC article.
-
Distinct Interaction Sites of Rac GTPase with WAVE Regulatory Complex Have Non-redundant Functions in Vivo.Curr Biol. 2018 Nov 19;28(22):3674-3684.e6. doi: 10.1016/j.cub.2018.10.002. Epub 2018 Nov 1. Curr Biol. 2018. PMID: 30393033 Free PMC article.
-
Structure and control of the actin regulatory WAVE complex.Nature. 2010 Nov 25;468(7323):533-8. doi: 10.1038/nature09623. Nature. 2010. PMID: 21107423 Free PMC article.
-
A role for the small GTPase Rac1 in vaccinia actin-based motility.Small GTPases. 2015;6(2):119-22. doi: 10.1080/21541248.2015.1055182. Small GTPases. 2015. PMID: 26147090 Free PMC article. Review.
-
Phosphoregulation of the WAVE regulatory complex and signal integration.Semin Cell Dev Biol. 2013 Apr;24(4):272-9. doi: 10.1016/j.semcdb.2013.01.007. Epub 2013 Jan 24. Semin Cell Dev Biol. 2013. PMID: 23354023 Free PMC article. Review.
Cited by
-
Potential therapeutic role of spermine via Rac1 in osteoporosis: Insights from zebrafish and mice.Zool Res. 2024 Mar 18;45(2):367-380. doi: 10.24272/j.issn.2095-8137.2023.371. Zool Res. 2024. PMID: 38485506 Free PMC article.
-
Oscillatory dynamics of Rac1 activity in Dictyostelium discoideum amoebae.PLoS Comput Biol. 2024 Dec 9;20(12):e1012025. doi: 10.1371/journal.pcbi.1012025. eCollection 2024 Dec. PLoS Comput Biol. 2024. PMID: 39652619 Free PMC article.
-
Arf GTPase activates the WAVE regulatory complex through a distinct binding site.Sci Adv. 2022 Dec 14;8(50):eadd1412. doi: 10.1126/sciadv.add1412. Epub 2022 Dec 14. Sci Adv. 2022. PMID: 36516255 Free PMC article.
-
Molecular basis of the CYFIP2 and NCKAP1 autism-linked variants in the WAVE regulatory complex.Protein Sci. 2025 Jan;34(1):e5238. doi: 10.1002/pro.5238. Protein Sci. 2025. PMID: 39660913 Free PMC article.
-
RAC1-Amplified and RAC1-A159V Hotspot-Mutated Head and Neck Cancer Sensitive to the Rac Inhibitor EHop-016 In Vivo: A Proof-of-Concept Study.Cancers (Basel). 2025 Jan 23;17(3):361. doi: 10.3390/cancers17030361. Cancers (Basel). 2025. PMID: 39941730 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
