

Reconstituted TAD-size chromatin fibers feature heterogeneous nucleosome clusters
- PMID: 36114220
- PMCID: PMC9481575
- DOI: 10.1038/s41598-022-19471-3
Reconstituted TAD-size chromatin fibers feature heterogeneous nucleosome clusters
Abstract
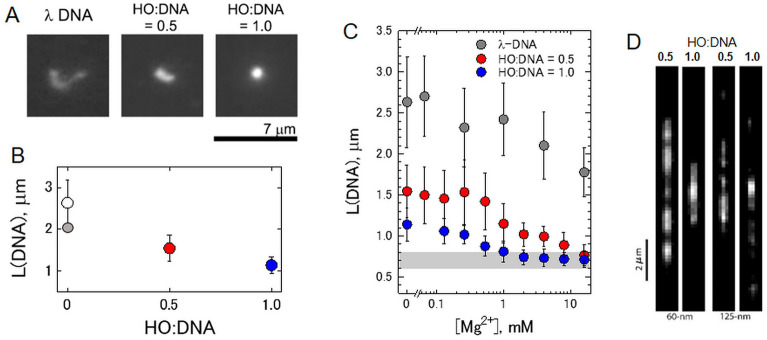
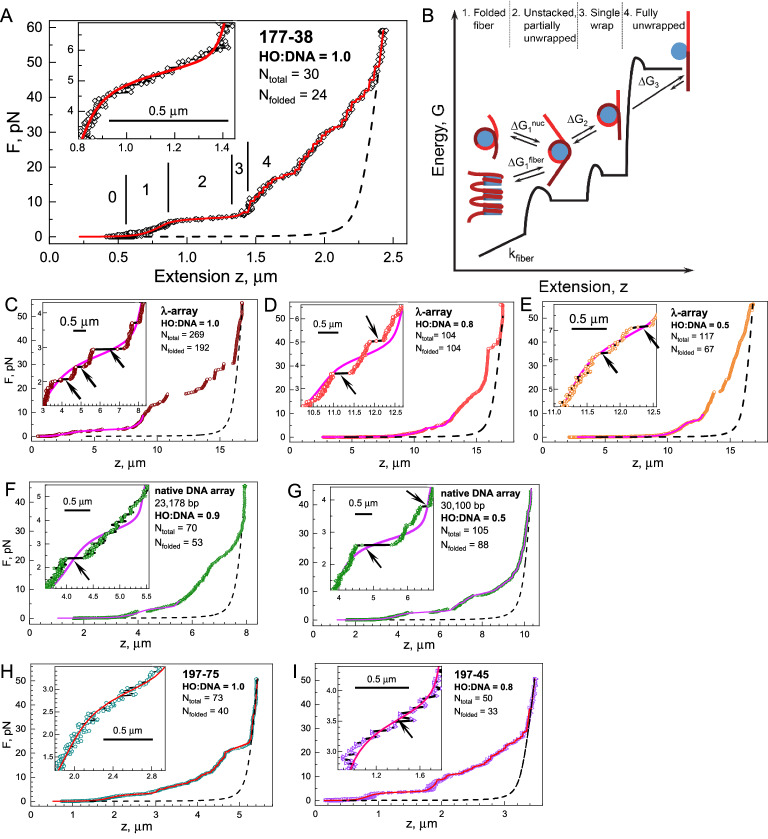
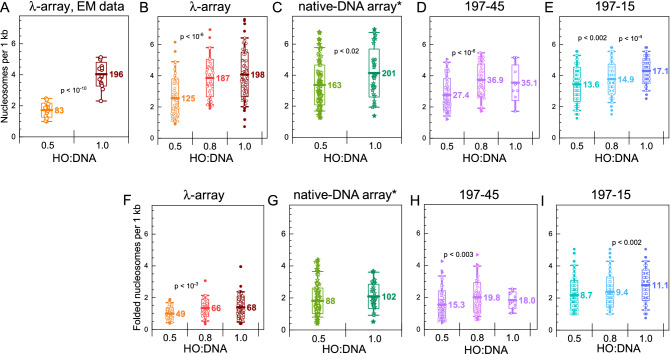
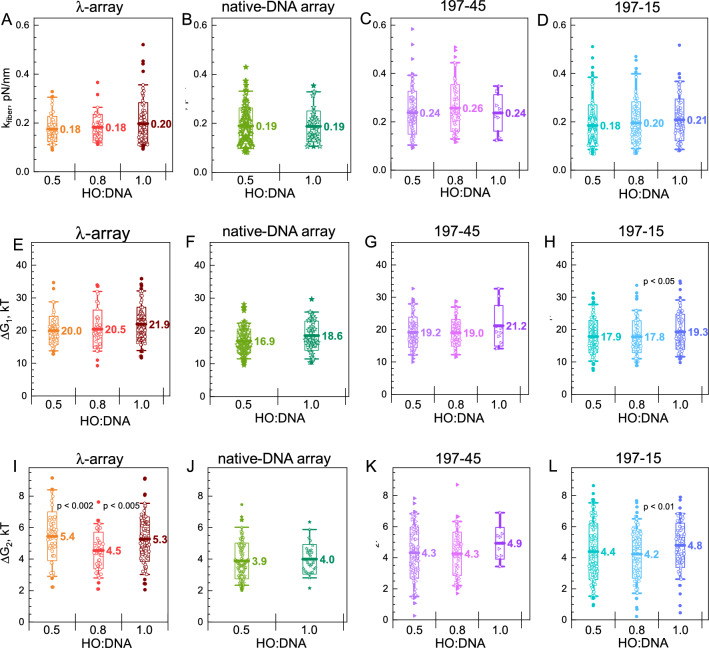
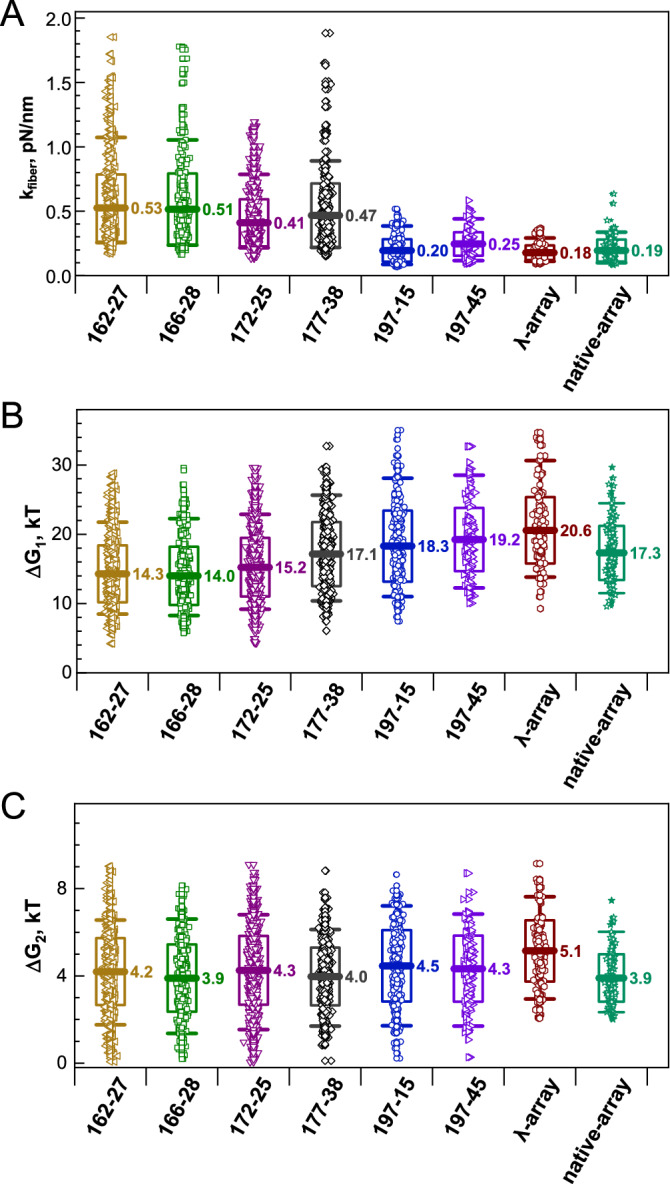
Large topologically associated domains (TADs) contain irregularly spaced nucleosome clutches, and interactions between such clutches are thought to aid the compaction of these domains. Here, we reconstituted TAD-sized chromatin fibers containing hundreds of nucleosomes on native source human and lambda-phage DNA and compared their mechanical properties at the single-molecule level with shorter '601' arrays with various nucleosome repeat lengths. Fluorescent imaging showed increased compaction upon saturation of the DNA with histones and increasing magnesium concentration. Nucleosome clusters and their structural fluctuations were visualized in confined nanochannels. Force spectroscopy revealed not only similar mechanical properties of the TAD-sized fibers as shorter fibers but also large rupture events, consistent with breaking the interactions between distant clutches of nucleosomes. Though the arrays of native human DNA, lambda-phage and '601' DNA featured minor differences in reconstitution yield and nucleosome stability, the fibers' global structural and mechanical properties were similar, including the interactions between nucleosome clutches. These single-molecule experiments quantify the mechanical forces that stabilize large TAD-sized chromatin domains consisting of disordered, dynamically interacting nucleosome clutches and their effect on the condensation of large chromatin domains.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Nucleosome Clutches are Regulated by Chromatin Internal Parameters.J Mol Biol. 2021 Mar 19;433(6):166701. doi: 10.1016/j.jmb.2020.11.001. Epub 2020 Nov 9. J Mol Biol. 2021. PMID: 33181171 Free PMC article.
-
Changing chromatin fiber conformation by nucleosome repositioning.Biophys J. 2014 Nov 4;107(9):2141-50. doi: 10.1016/j.bpj.2014.09.026. Biophys J. 2014. PMID: 25418099 Free PMC article.
-
Nucleosome spacing and chromatin higher-order folding.Nucleus. 2012 Nov-Dec;3(6):493-9. doi: 10.4161/nucl.22168. Epub 2012 Sep 18. Nucleus. 2012. PMID: 22990522 Free PMC article.
-
Breaths, Twists, and Turns of Atomistic Nucleosomes.J Mol Biol. 2021 Mar 19;433(6):166744. doi: 10.1016/j.jmb.2020.166744. Epub 2020 Dec 10. J Mol Biol. 2021. PMID: 33309853 Review.
-
Nucleosome positioning and chromatin organization.Curr Opin Struct Biol. 2020 Oct;64:111-118. doi: 10.1016/j.sbi.2020.06.021. Epub 2020 Jul 27. Curr Opin Struct Biol. 2020. PMID: 32731156 Review.
Cited by
-
Genome organization across scales: mechanistic insights from in vitro reconstitution studies.Biochem Soc Trans. 2024 Apr 24;52(2):793-802. doi: 10.1042/BST20230883. Biochem Soc Trans. 2024. PMID: 38451192 Free PMC article. Review.
-
The shelterin component TRF2 mediates columnar stacking of human telomeric chromatin.EMBO J. 2024 Jan;43(1):87-111. doi: 10.1038/s44318-023-00002-3. Epub 2023 Dec 14. EMBO J. 2024. PMID: 38177309 Free PMC article.
-
Dynamics of nucleosomes and chromatin fibers revealed by single-molecule measurements.BMB Rep. 2025 Jan;58(1):24-32. doi: 10.5483/BMBRep.2024-0191. BMB Rep. 2025. PMID: 39757199 Free PMC article. Review.
-
Chromatin Liquid-Liquid Phase Separation (LLPS) Is Regulated by Ionic Conditions and Fiber Length.Cells. 2022 Oct 6;11(19):3145. doi: 10.3390/cells11193145. Cells. 2022. PMID: 36231107 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
