

Pentacyclic triterpenoid ursolic acid interferes with mast cell activation via a lipid-centric mechanism affecting FcεRI signalosome functions
- PMID: 36115460
- PMCID: PMC9587013
- DOI: 10.1016/j.jbc.2022.102497
Pentacyclic triterpenoid ursolic acid interferes with mast cell activation via a lipid-centric mechanism affecting FcεRI signalosome functions
Abstract
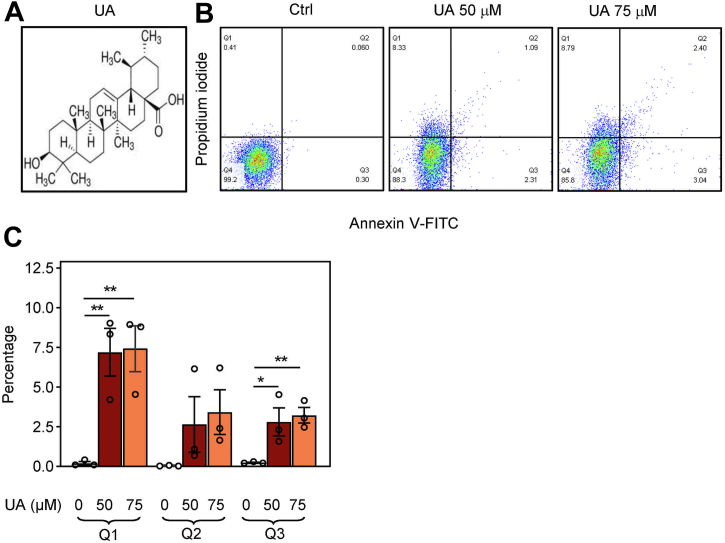
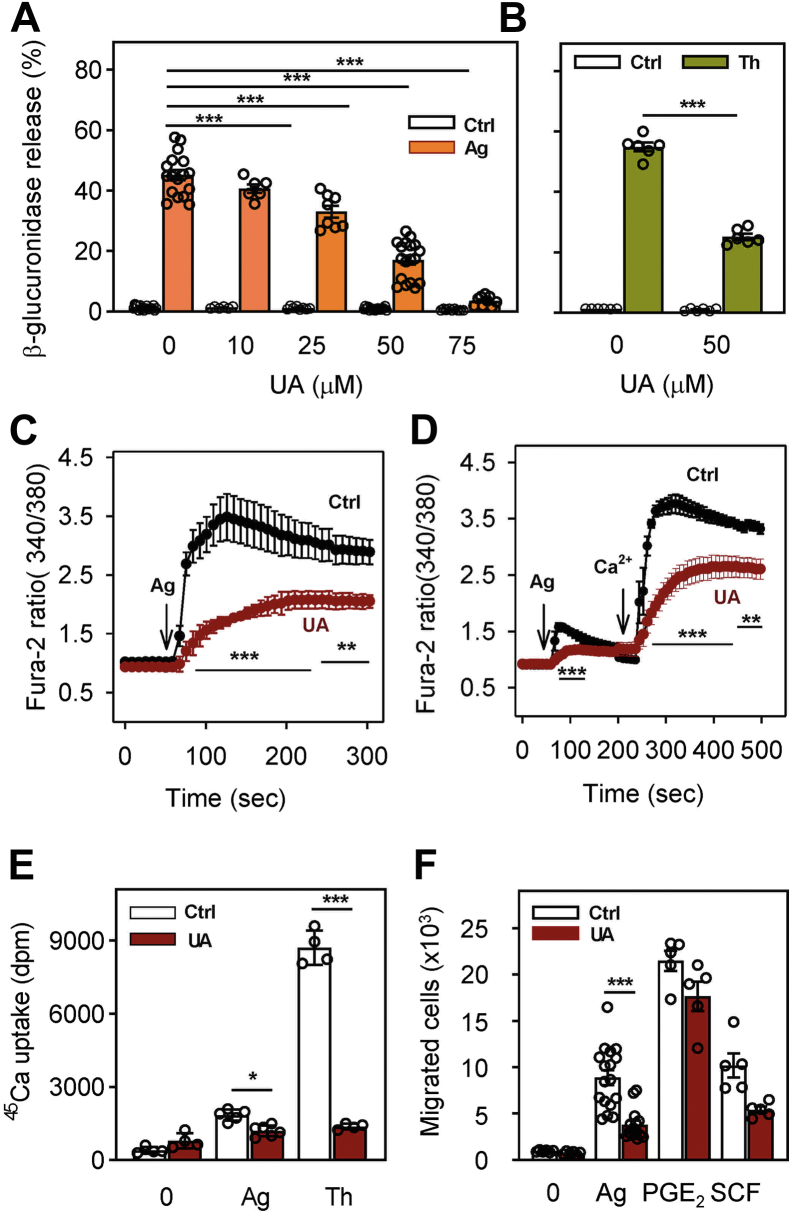
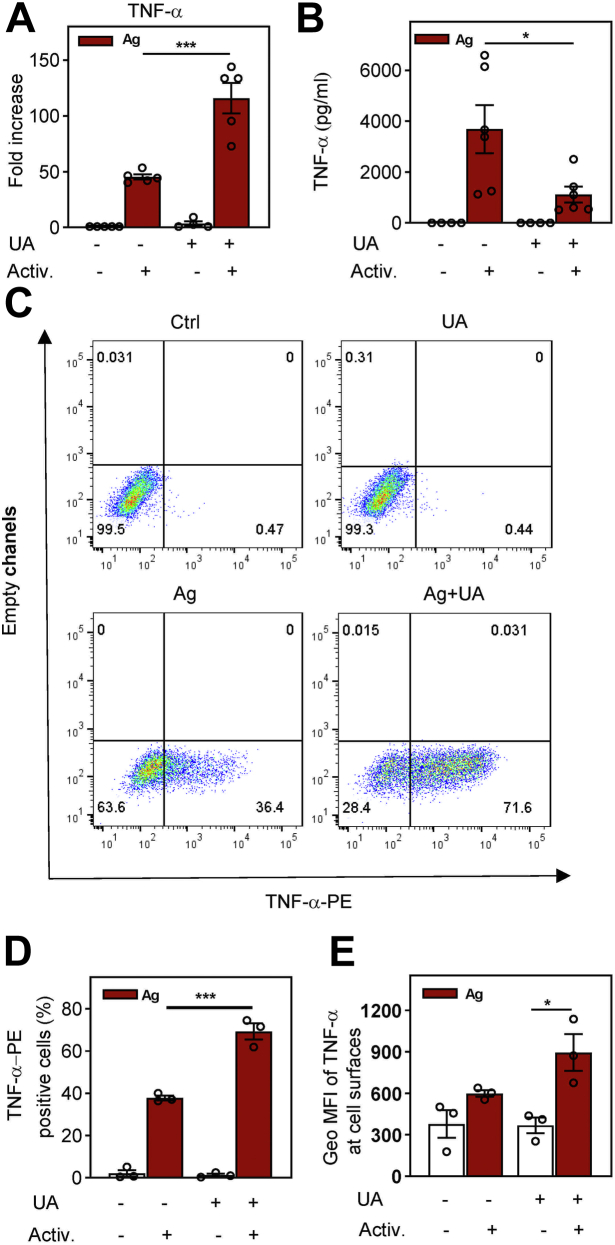
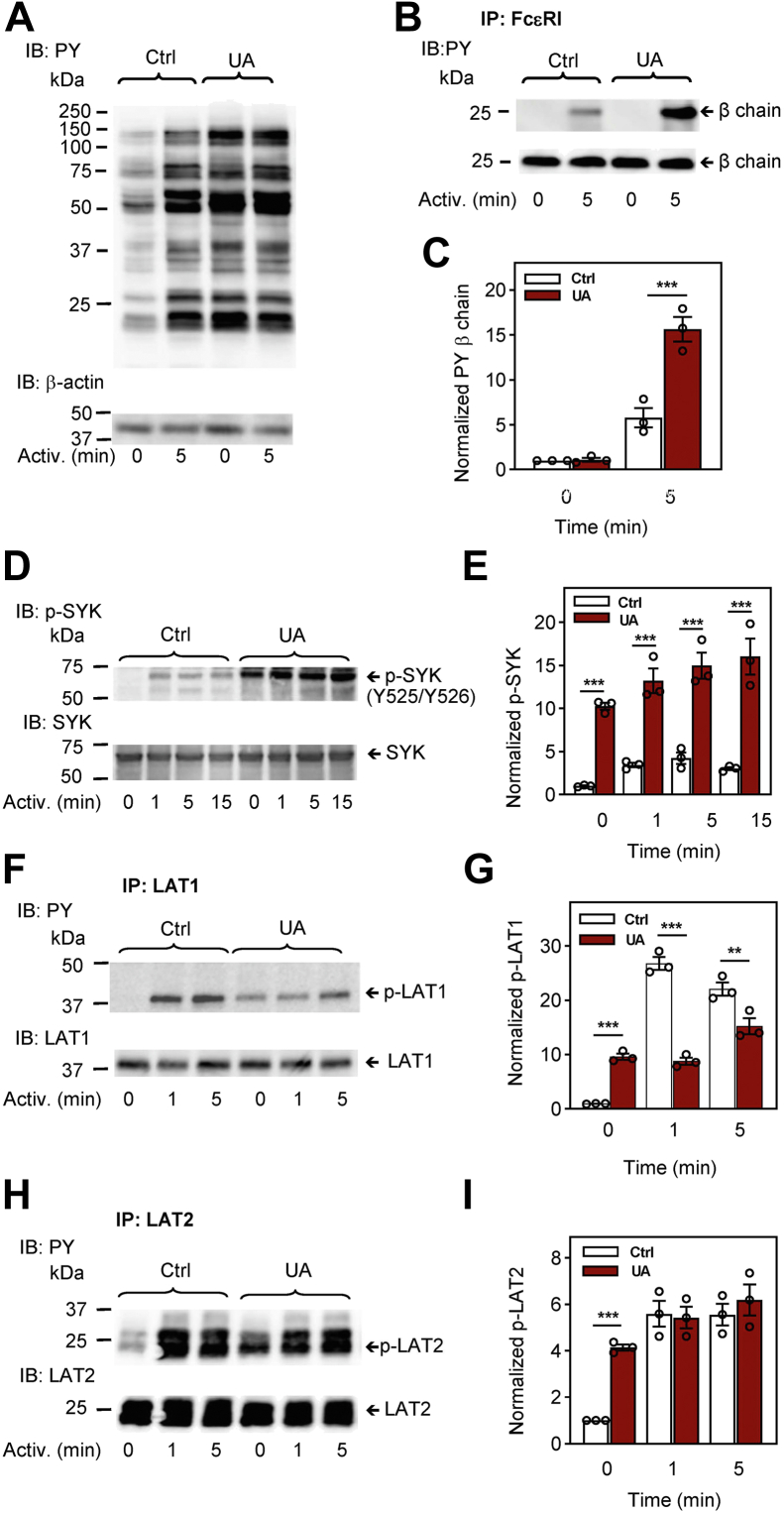
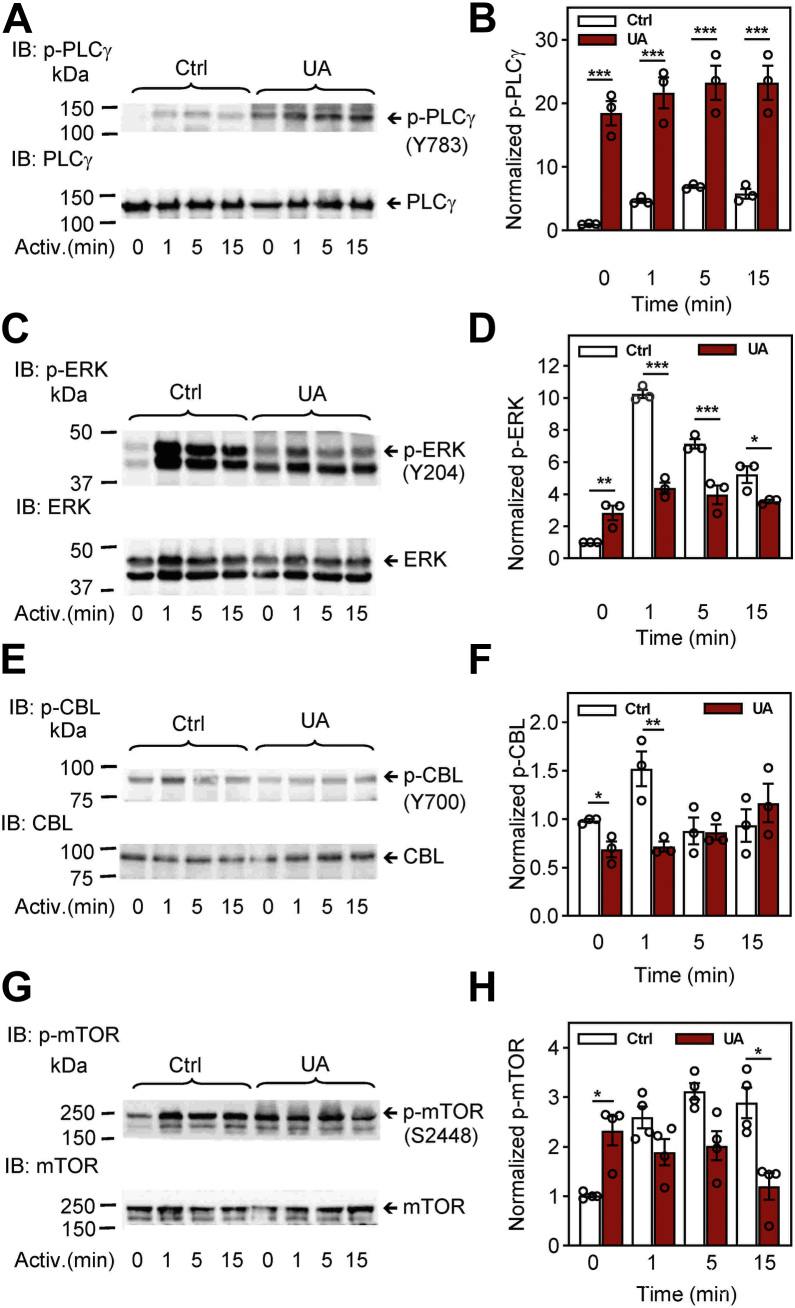
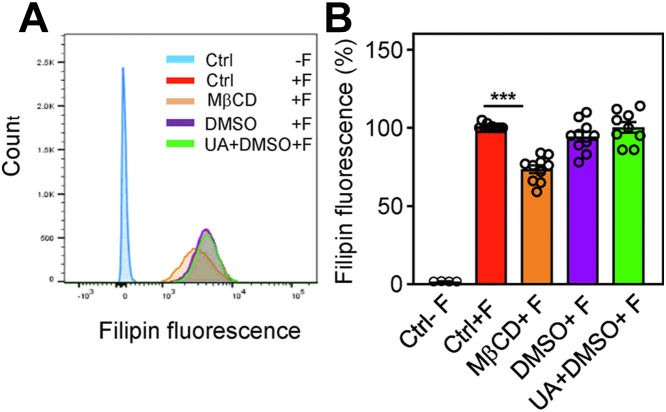
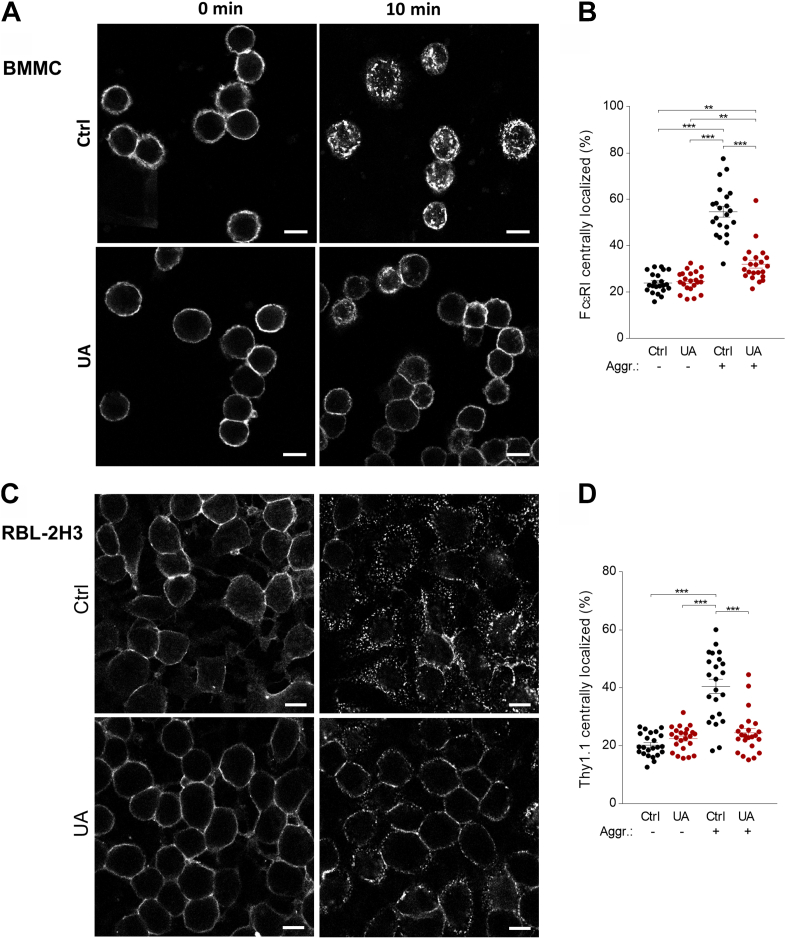
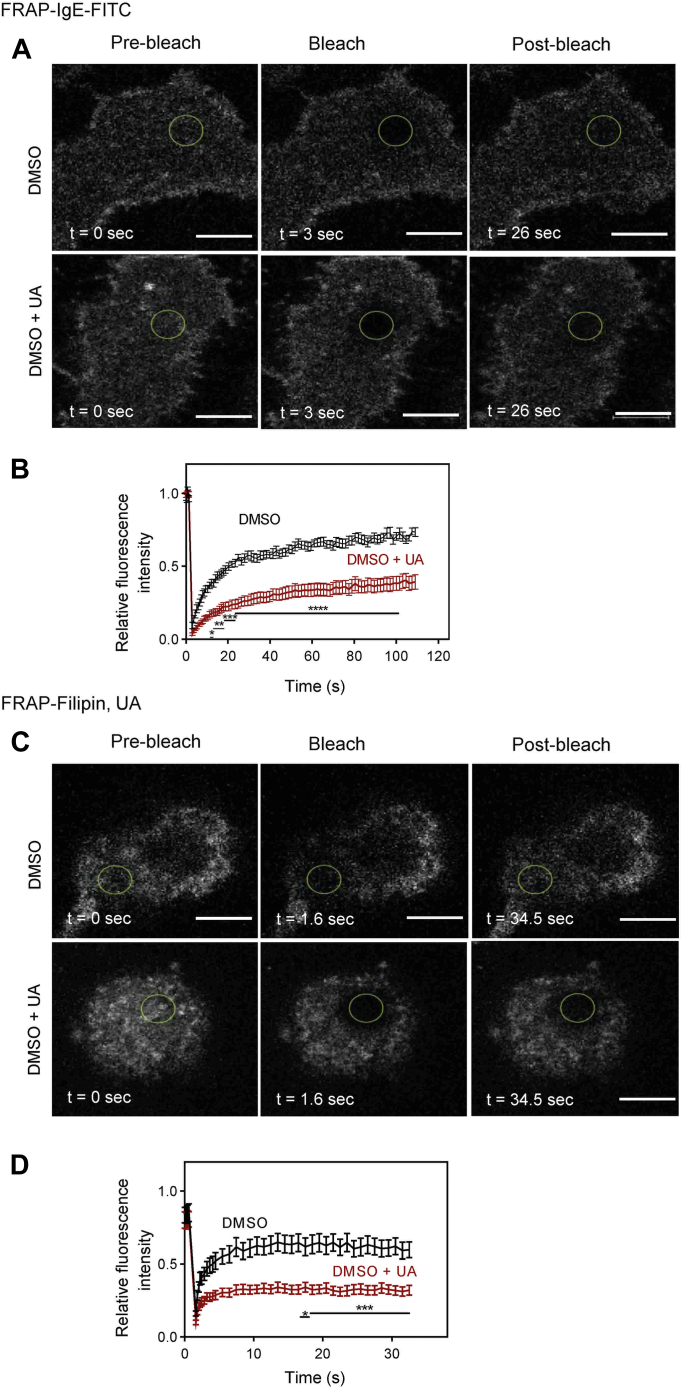
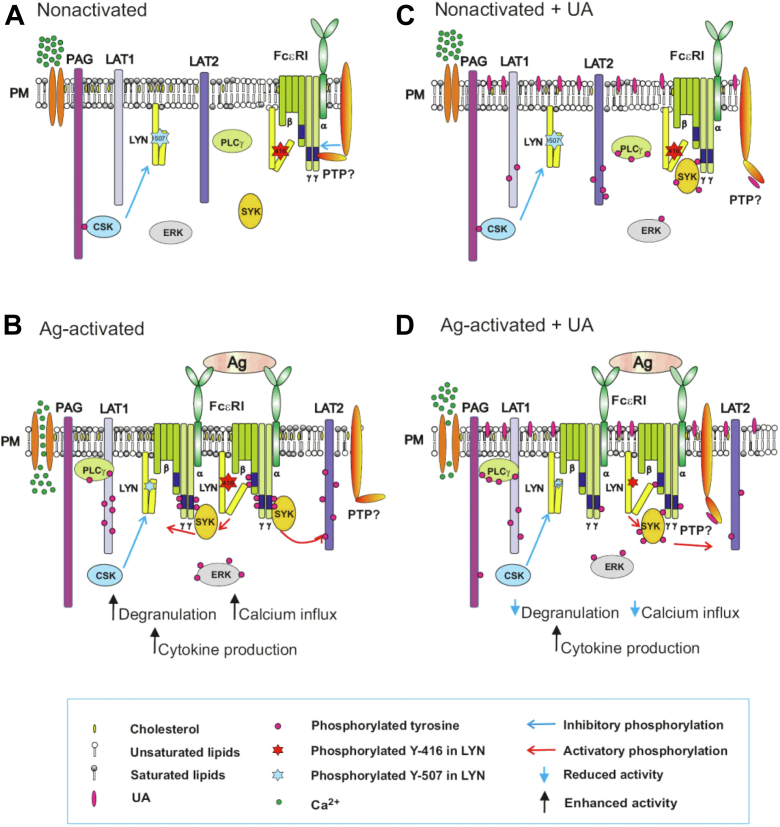
Pentacyclic triterpenoids, including ursolic acid (UA), are bioactive compounds with multiple biological activities involving anti-inflammatory effects. However, the mode of their action on mast cells, key players in the early stages of allergic inflammation, and underlying molecular mechanisms remain enigmatic. To better understand the effect of UA on mast cell signaling, here we examined the consequences of short-term treatment of mouse bone marrow-derived mast cells with UA. Using IgE-sensitized and antigen- or thapsigargin-activated cells, we found that 15 min exposure to UA inhibited high affinity IgE receptor (FcεRI)-mediated degranulation, calcium response, and extracellular calcium uptake. We also found that UA inhibited migration of mouse bone marrow-derived mast cells toward antigen but not toward prostaglandin E2 and stem cell factor. Compared to control antigen-activated cells, UA enhanced the production of tumor necrosis factor-α at the mRNA and protein levels. However, secretion of this cytokine was inhibited. Further analysis showed that UA enhanced tyrosine phosphorylation of the SYK kinase and several other proteins involved in the early stages of FcεRI signaling, even in the absence of antigen activation, but inhibited or reduced their further phosphorylation at later stages. In addition, we show that UA induced changes in the properties of detergent-resistant plasma membrane microdomains and reduced antibody-mediated clustering of the FcεRI and glycosylphosphatidylinositol-anchored protein Thy-1. Finally, UA inhibited mobility of the FcεRI and cholesterol. These combined data suggest that UA exerts its effects, at least in part, via lipid-centric plasma membrane perturbations, hence affecting the functions of the FcεRI signalosome.
Keywords: immunoglobulin E; lipid raft; mast cell; plasma membrane; signal transduction; tumor necrosis factor; tyrosine kinase.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare that they have no conflict of interest with the contents of this article.
Figures
References
-
- Rivera J., Gilfillan A.M. Molecular regulation of mast cell activation. J. Allergy Clin. Immunol. 2006;117:1214–1225. - PubMed
-
- Harvima I.T., Levi-Schaffer F., Draber P., Friedman S., Polakovicova I., Gibbs B.F., et al. Molecular targets on mast cells and basophils for novel therapies. J. Allergy Clin. Immunol. 2014;134:530–544. - PubMed
-
- Halova I., Ronnberg E., Draberova L., Vliagoftis H., Nilsson G.P., Draber P. Changing the threshold - signals and mechanisms of mast cell priming. Immunol. Rev. 2018;282:73–86. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
