

The central extended amygdala guides survival-relevant tradeoffs: Implications for understanding common psychiatric disorders
- PMID: 36115597
- PMCID: PMC11178236
- DOI: 10.1016/j.neubiorev.2022.104879
The central extended amygdala guides survival-relevant tradeoffs: Implications for understanding common psychiatric disorders
Abstract
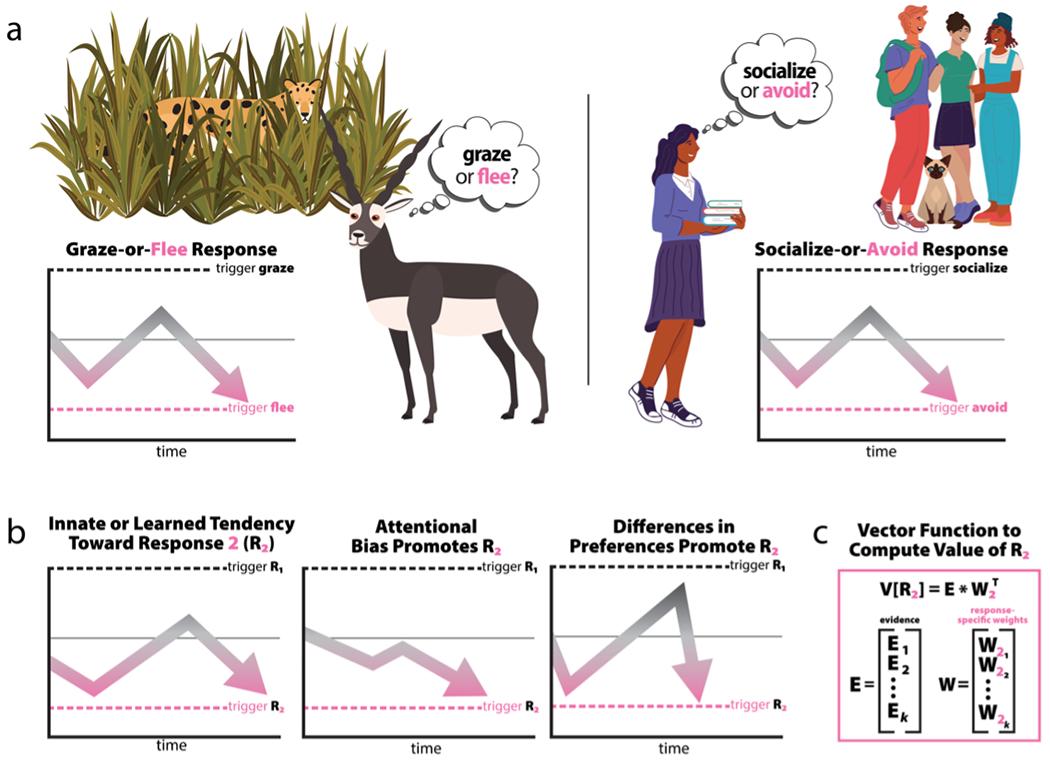
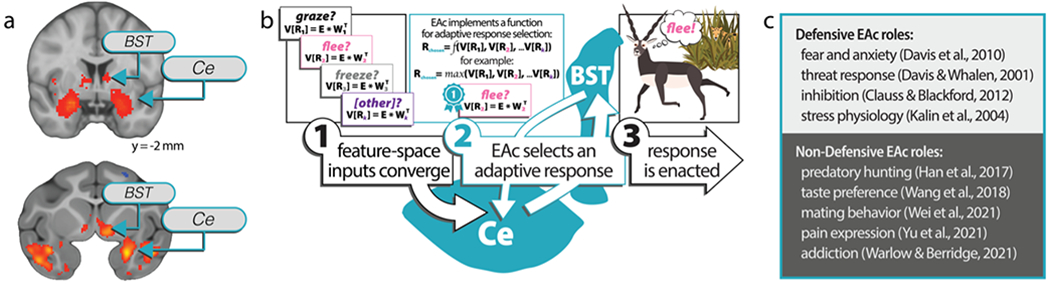
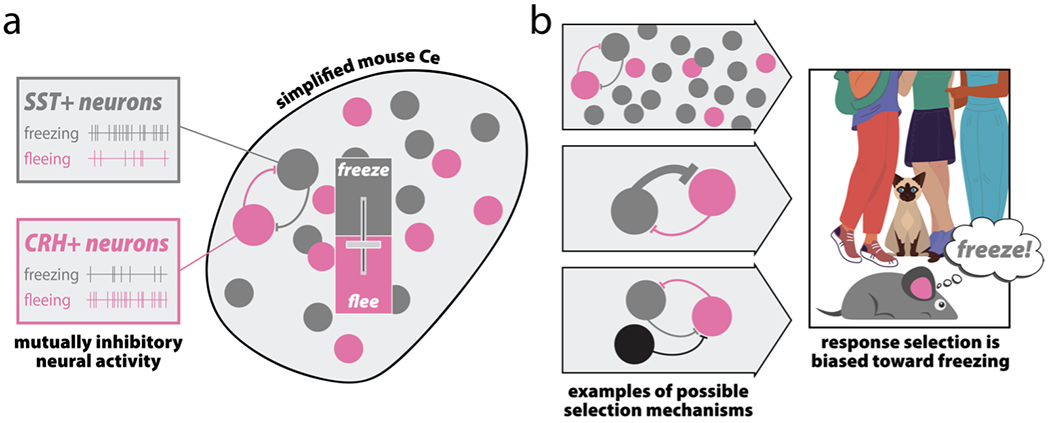
To thrive in challenging environments, individuals must pursue rewards while avoiding threats. Extensive studies in animals and humans have identified the central extended amygdala (EAc)-which includes the central nucleus of the amygdala (Ce) and bed nucleus of the stria terminalis (BST)-as a conserved substrate for defensive behavior. These studies suggest the EAc influences defensive responding and assembles fearful and anxious states. This has led to the proliferation of a view that the EAc is fundamentally a defensive substrate. Yet mechanistic work in animals has implicated the EAc in numerous appetitive and consummatory processes, yielding fresh insights into the microcircuitry of survival- and emotion-relevant response selection. Coupled with the EAc's centrality in a conserved network of brain regions that encode multisensory environmental and interoceptive information, these findings suggest a broader role for the EAc as an arbiter of survival- and emotion-relevant tradeoffs for action selection. Determining how the EAc optimizes these tradeoffs promises to improve our understanding of common psychiatric illnesses such as anxiety, depression, alcohol- and substance-use disorders, and anhedonia.
Keywords: Action selection; Anxiety; BNST; BST; Bed nucleus of the stria terminalis; Ce; CeA; Central nucleus of the amygdala; Consummatory; Defensive behavior; Extended amygdala; Fear; Reward.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Figures
References
-
- Aggleton JP (2000). The amygdala: A functional analysis. Oxford University Press, Oxford, UK.
