

TGF-β signaling in the tumor metabolic microenvironment and targeted therapies
- PMID: 36115986
- PMCID: PMC9482317
- DOI: 10.1186/s13045-022-01349-6
TGF-β signaling in the tumor metabolic microenvironment and targeted therapies
Abstract
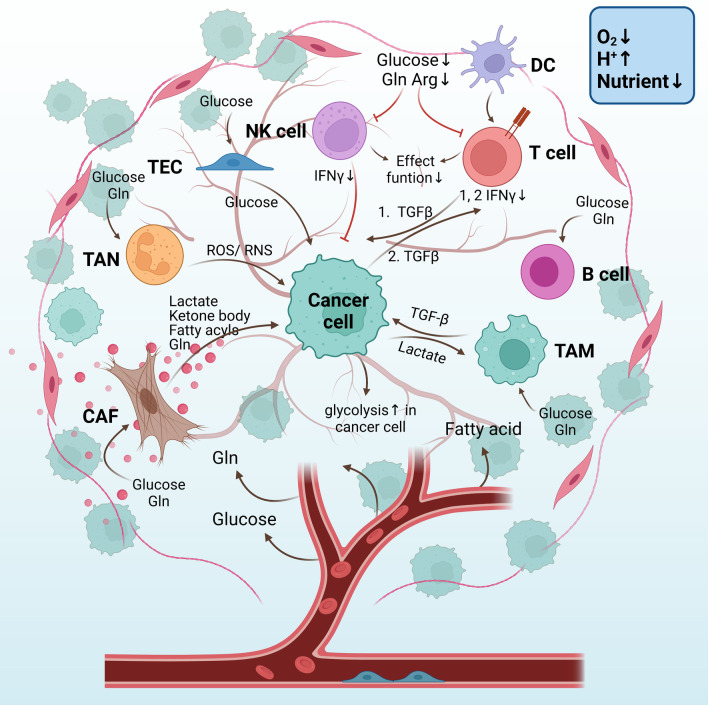
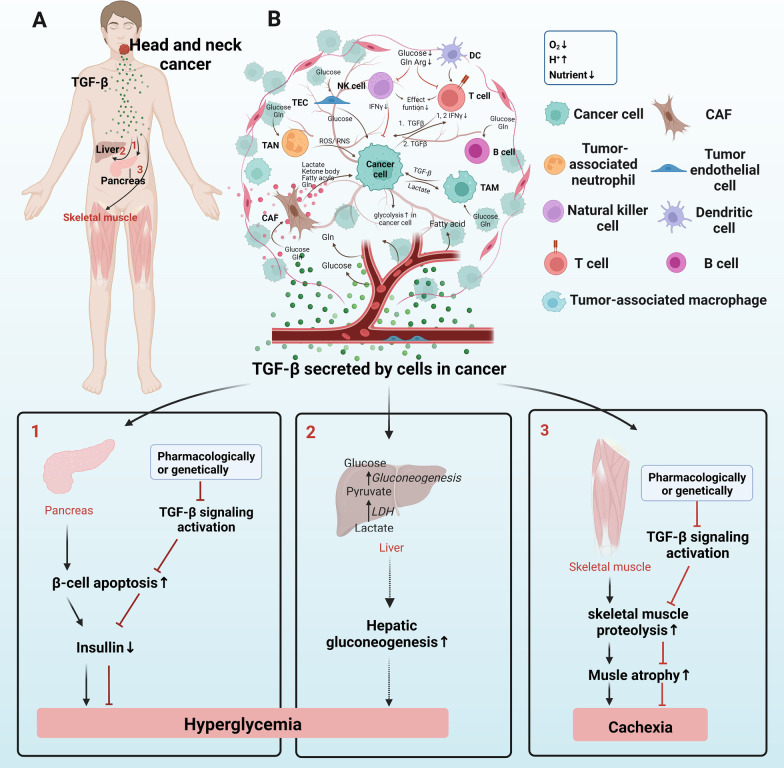
Transforming growth factor-β (TGF-β) signaling has a paradoxical role in cancer progression, and it acts as a tumor suppressor in the early stages but a tumor promoter in the late stages of cancer. Once cancer cells are generated, TGF-β signaling is responsible for the orchestration of the immunosuppressive tumor microenvironment (TME) and supports cancer growth, invasion, metastasis, recurrence, and therapy resistance. These progressive behaviors are driven by an "engine" of the metabolic reprogramming in cancer. Recent studies have revealed that TGF-β signaling regulates cancer metabolic reprogramming and is a metabolic driver in the tumor metabolic microenvironment (TMME). Intriguingly, TGF-β ligands act as an "endocrine" cytokine and influence host metabolism. Therefore, having insight into the role of TGF-β signaling in the TMME is instrumental for acknowledging its wide range of effects and designing new cancer treatment strategies. Herein, we try to illustrate the concise definition of TMME based on the published literature. Then, we review the metabolic reprogramming in the TMME and elaborate on the contribution of TGF-β to metabolic rewiring at the cellular (intracellular), tissular (intercellular), and organismal (cancer-host) levels. Furthermore, we propose three potential applications of targeting TGF-β-dependent mechanism reprogramming, paving the way for TGF-β-related antitumor therapy from the perspective of metabolism.
Keywords: Cancer cell; Host metabolism; Stromal cell; TGF-β signaling; Tumor metabolic microenvironment.
© 2022. The Author(s).
Conflict of interest statement
All other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
