

Role of cathepsin B of Naegleria fowleri during primary amebic meningoencephalitis
- PMID: 36125528
- PMCID: PMC9485797
- DOI: 10.1007/s00436-022-07660-y
Role of cathepsin B of Naegleria fowleri during primary amebic meningoencephalitis
Abstract
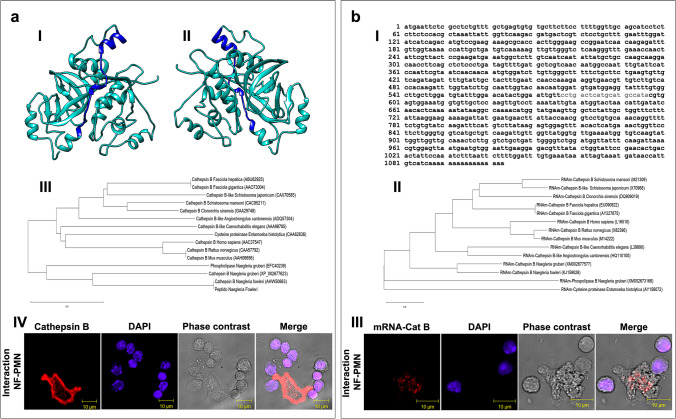
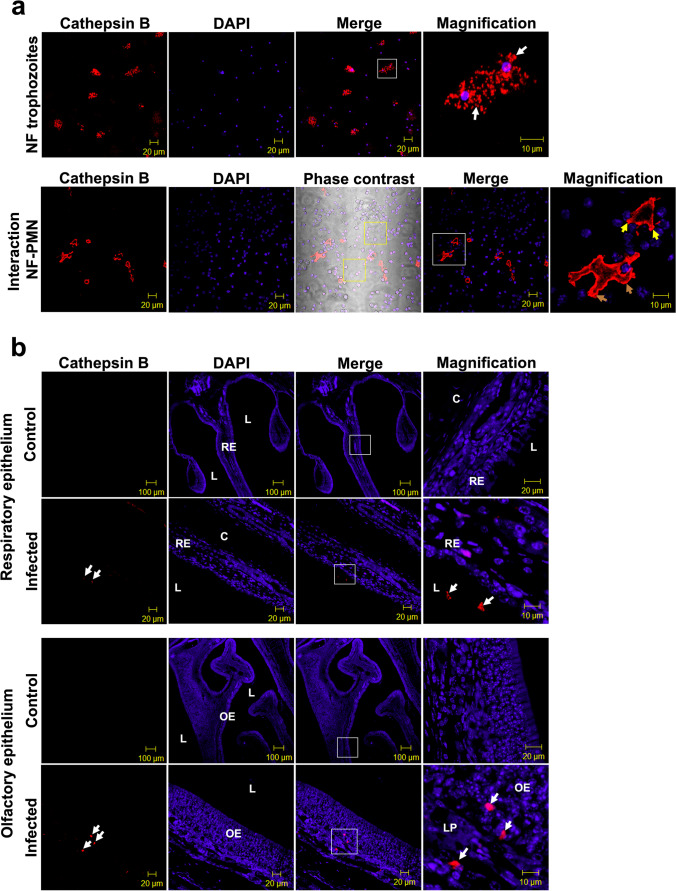
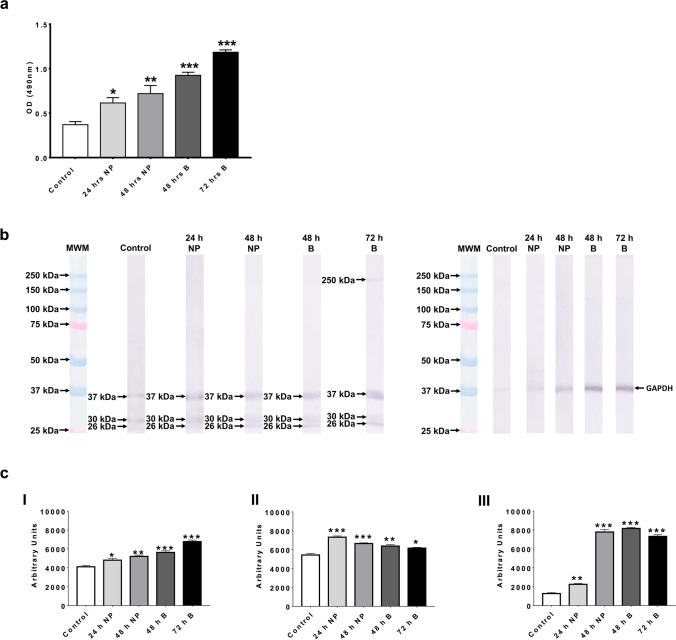
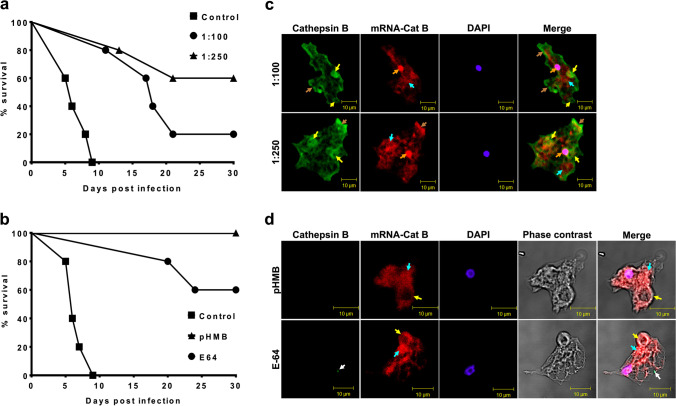
Naegleria fowleri causes primary amoebic meningoencephalitis in humans and experimental animals. It has been suggested that cysteine proteases of parasites play key roles in metabolism, nutrient uptake, host tissue invasion, and immune evasion. The aim of this work was to evaluate the presence, expression, and role of cathepsin B from N. fowleri in vitro and during PAM. Rabbit-specific polyclonal antibodies against cathepsin B were obtained from rabbit immunization with a synthetic peptide obtained by bioinformatic design. In addition, a probe was designed from mRNA for N. fowleri cathepsin B. Both protein and messenger were detected in fixed trophozoites, trophozoites interacted with polymorphonuclear and histological sections of infected mice. The main cathepsin B distribution was observed in cytoplasm or membrane mainly pseudopods and food-cups while messenger was in nucleus and cytoplasm. Surprisingly, both the messenger and enzyme were observed in extracellular medium. To determine cathepsin B release, we used trophozoites supernatant recovered from nasal passages or brain of infected mice. We observed the highest release in supernatant from recovered brain amoebae, and when we analyzed molecular weight of secreted proteins by immunoblot, we found 30 and 37 kDa bands which were highly immunogenic. Finally, role of cathepsin B during N. fowleri infection was determined; we preincubated trophozoites with E-64, pHMB or antibodies with which we obtained 60%, 100%, and 60% of survival, respectively, in infected mice. These results suggest that cathepsin B plays a role during pathogenesis caused by N. fowleri mainly in adhesion and contributes to nervous tissue damage.
Keywords: Naegleria fowleri; antibodies; cathepsin B; cysteine proteases; protease inhibitors.
© 2022. The Author(s), under exclusive licence to Springer-Verlag GmbH Germany, part of Springer Nature.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
