

Sleep exerts lasting effects on hematopoietic stem cell function and diversity
- PMID: 36129517
- PMCID: PMC9499822
- DOI: 10.1084/jem.20220081
Sleep exerts lasting effects on hematopoietic stem cell function and diversity
Abstract
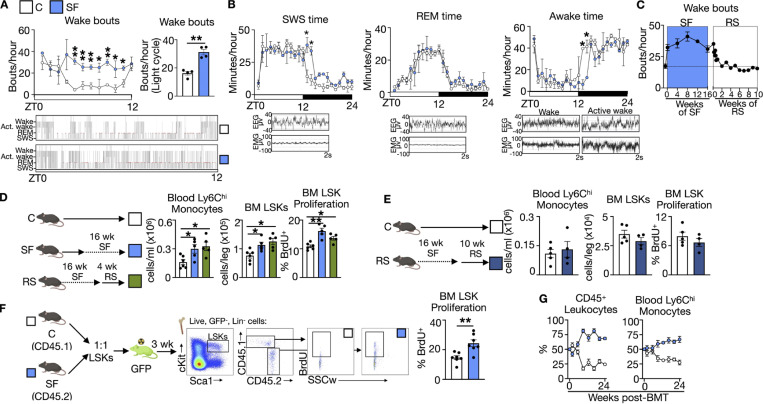
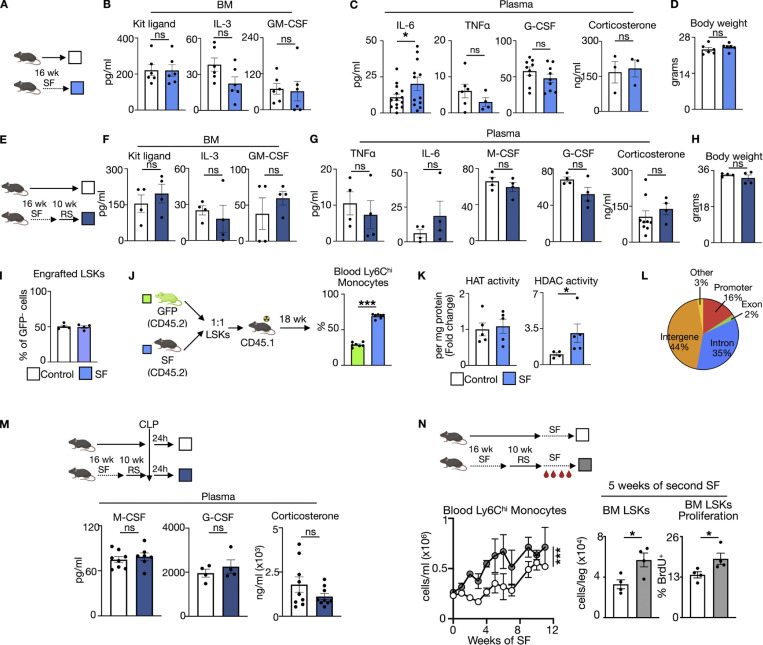
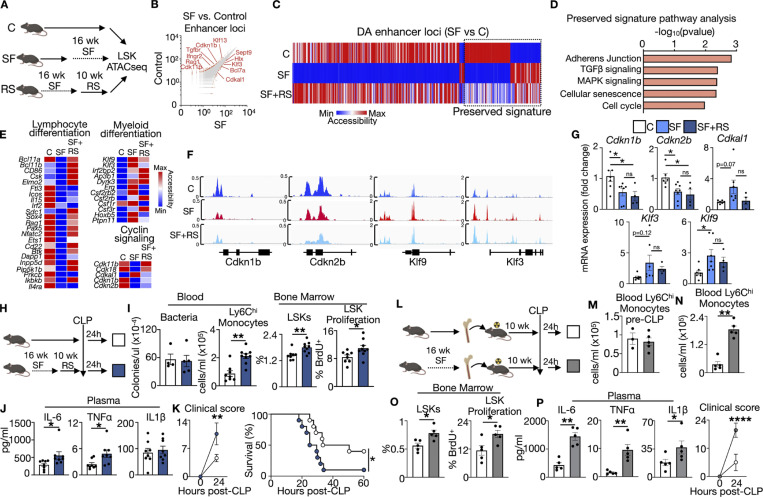
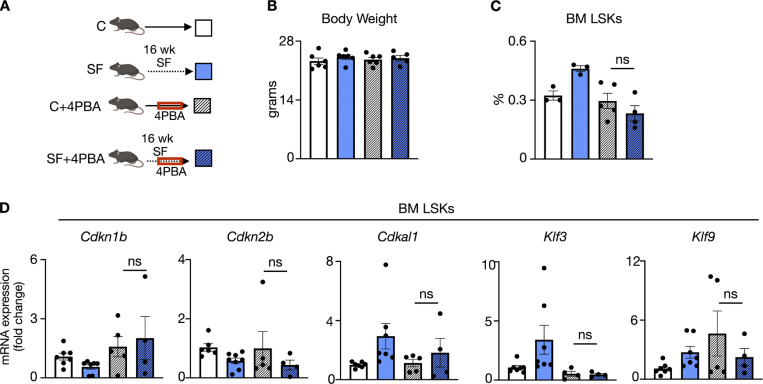
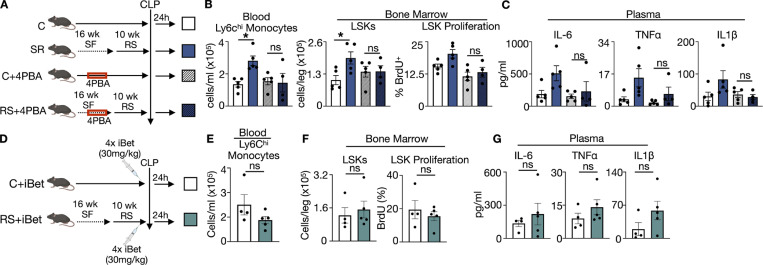
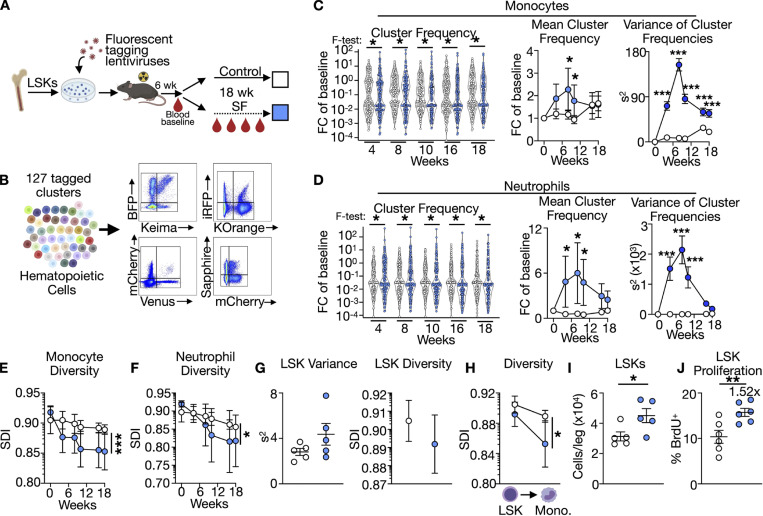
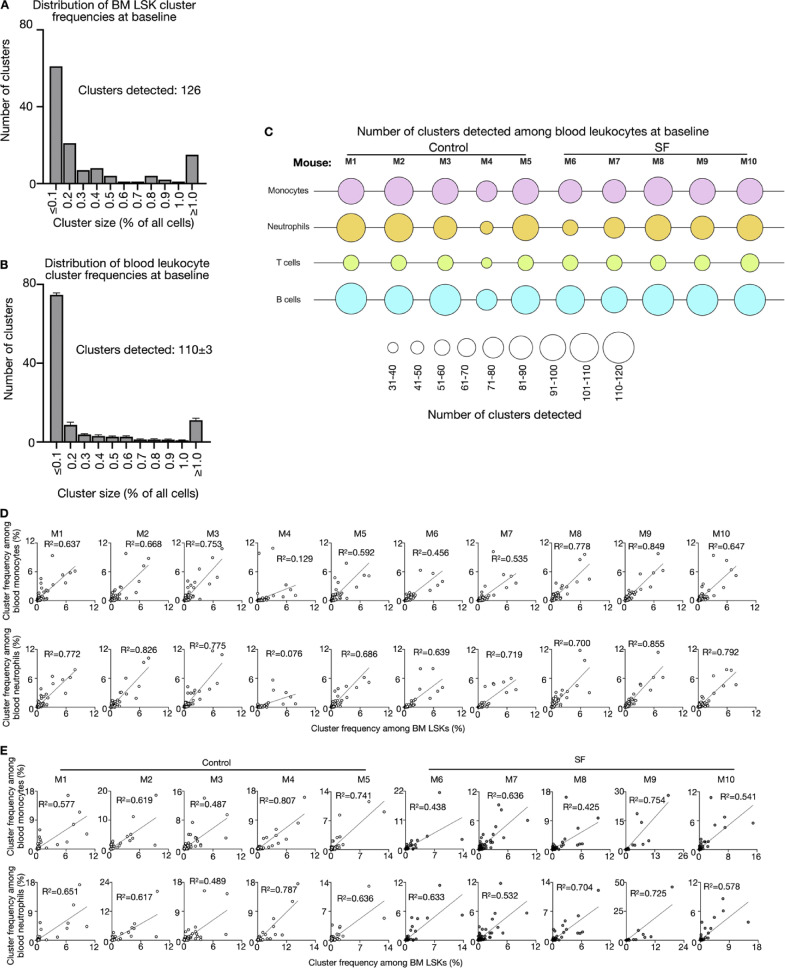
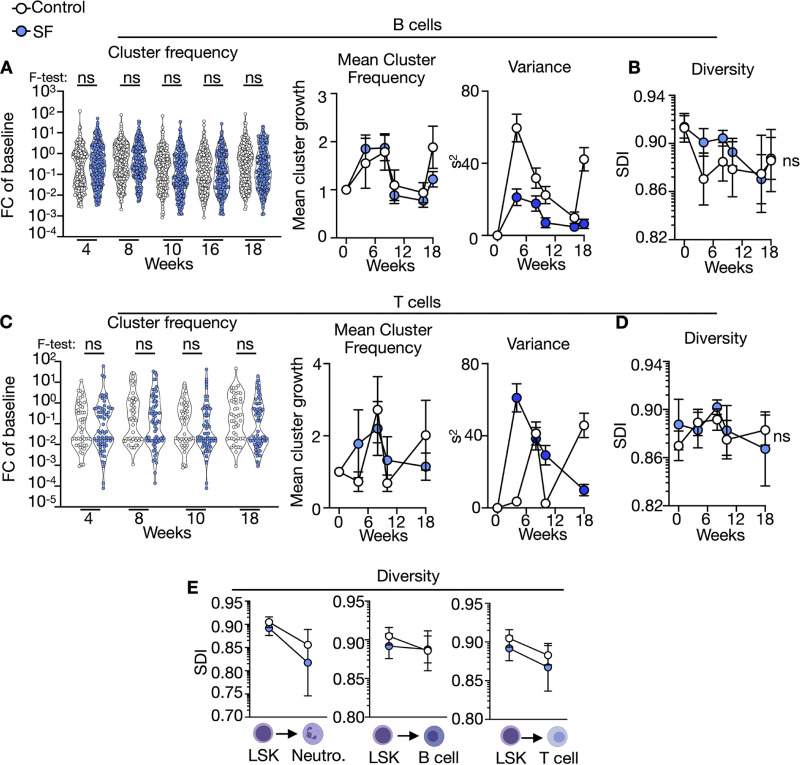
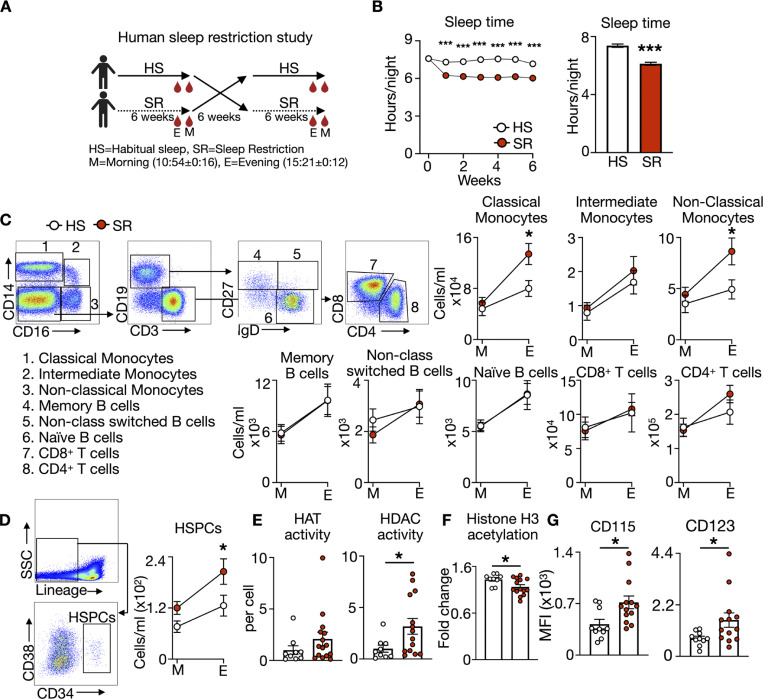
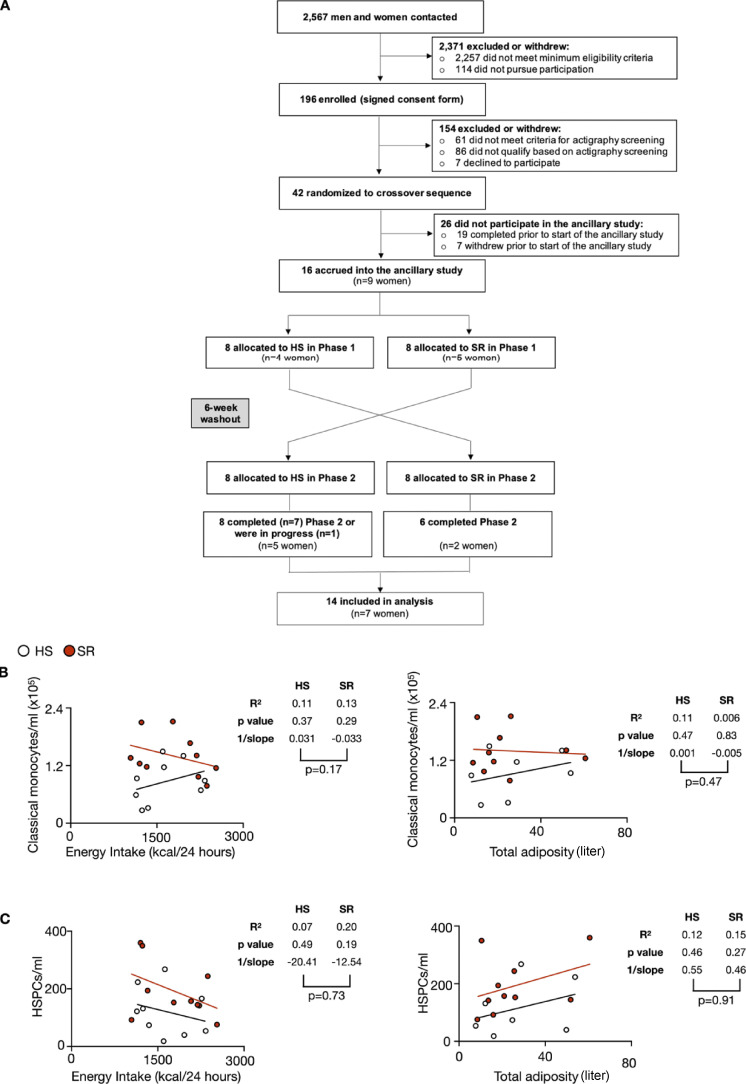
A sleepless night may feel awful in its aftermath, but sleep's revitalizing powers are substantial, perpetuating the idea that convalescent sleep is a consequence-free physiological reset. Although recent studies have shown that catch-up sleep insufficiently neutralizes the negative effects of sleep debt, the mechanisms that control prolonged effects of sleep disruption are not understood. Here, we show that sleep interruption restructures the epigenome of hematopoietic stem and progenitor cells (HSPCs) and increases their proliferation, thus reducing hematopoietic clonal diversity through accelerated genetic drift. Sleep fragmentation exerts a lasting influence on the HSPC epigenome, skewing commitment toward a myeloid fate and priming cells for exaggerated inflammatory bursts. Combining hematopoietic clonal tracking with mathematical modeling, we infer that sleep preserves clonal diversity by limiting neutral drift. In humans, sleep restriction alters the HSPC epigenome and activates hematopoiesis. These findings show that sleep slows decay of the hematopoietic system by calibrating the hematopoietic epigenome, constraining inflammatory output, and maintaining clonal diversity.
© 2022 McAlpine et al.
Conflict of interest statement
Disclosures: M. Nahrendorf reported receiving funds or material research support from Lilly, Alnylam, Biotronik, CSL Behring, GlycoMimetics, GSK, Medtronic, Novartis, and Pfizer, as well as consulting fees from Biogen, Gimv, IFM Therapeutics, Molecular Imaging, Sigilon, NovoNordisk, Verseau Therapeutics, and Bitterroot. No other disclosures were reported.
Figures
References
-
- Cedernaes, J., Schonke M., Westholm J.O., Mi J., Chibalin A., Voisin S., Osler M., Vogel H., Hornaeus K., Dickson S.L., et al. . 2018. Acute sleep loss results in tissue-specific alterations in genome-wide DNA methylation state and metabolic fuel utilization in humans. Sci. Adv. 4:eaar8590. 10.1126/sciadv.aar8590 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
- P30 DK040561/DK/NIDDK NIH HHS/United States
- R01 HL158534/HL/NHLBI NIH HHS/United States
- P01 HL131478/HL/NHLBI NIH HHS/United States
- K99 HL151750/HL/NHLBI NIH HHS/United States
- T32 HL007343/HL/NHLBI NIH HHS/United States
- R35 HL139598/HL/NHLBI NIH HHS/United States
- R35 HL155670/HL/NHLBI NIH HHS/United States
- R00 HL151750/HL/NHLBI NIH HHS/United States
- R35 HL135752/HL/NHLBI NIH HHS/United States
- F31 HL147364/HL/NHLBI NIH HHS/United States
- P01 HL142494/HL/NHLBI NIH HHS/United States
- R01 HL128226/HL/NHLBI NIH HHS/United States
- R01 HL142648/HL/NHLBI NIH HHS/United States
- UL1 TR001873/TR/NCATS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
