

Vivaxin genes encode highly immunogenic, non-variant antigens on the Trypanosoma vivax cell-surface
- PMID: 36129968
- PMCID: PMC9529106
- DOI: 10.1371/journal.pntd.0010791
Vivaxin genes encode highly immunogenic, non-variant antigens on the Trypanosoma vivax cell-surface
Abstract
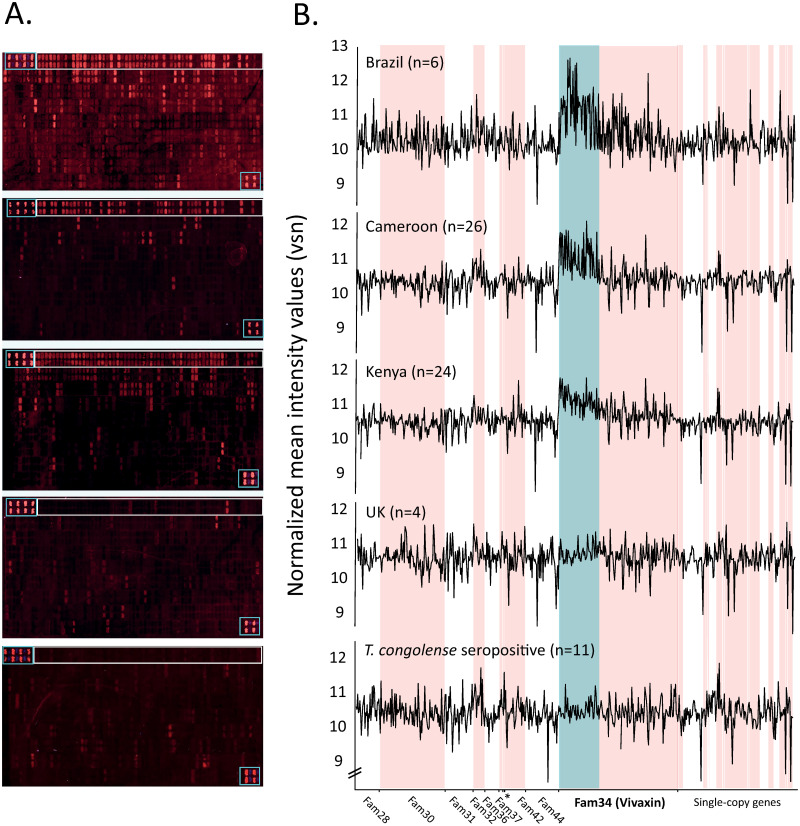
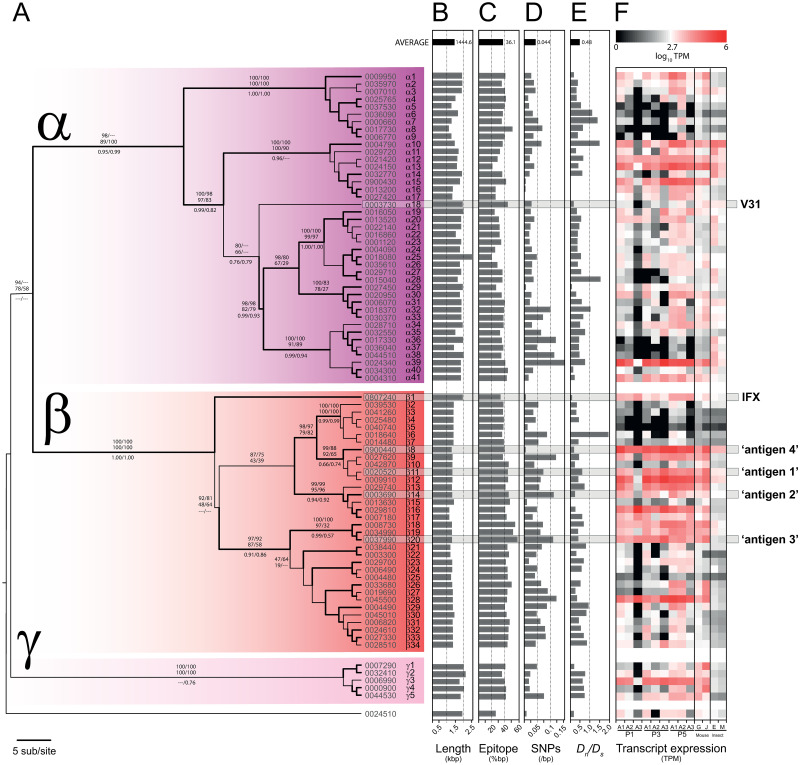
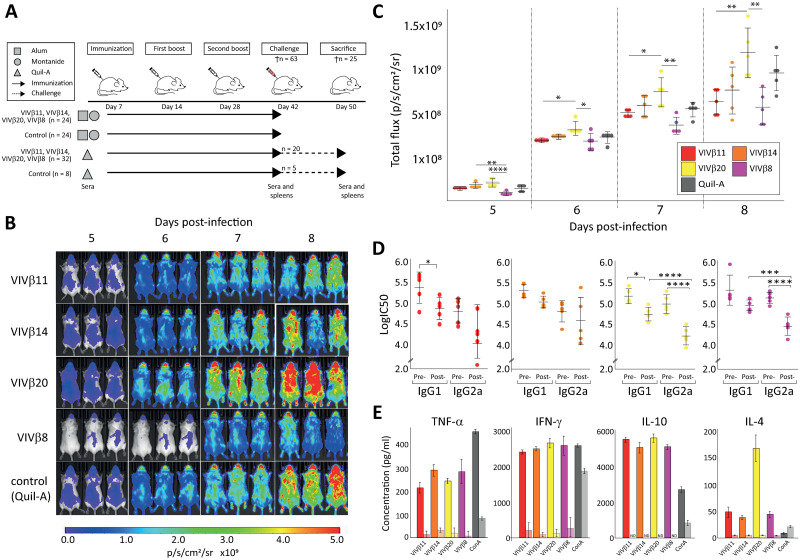
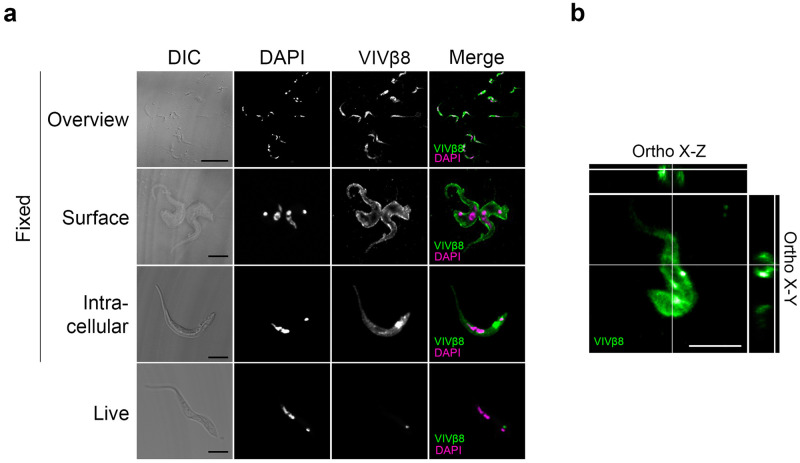
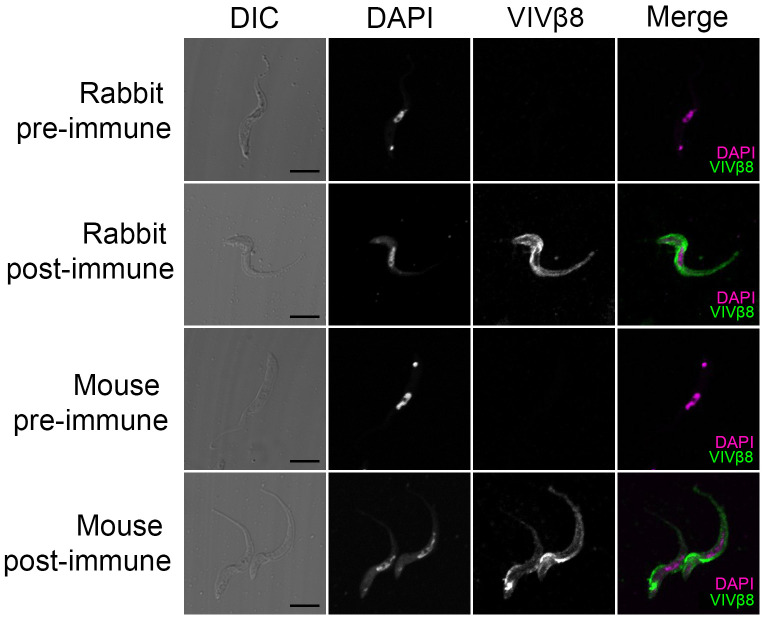
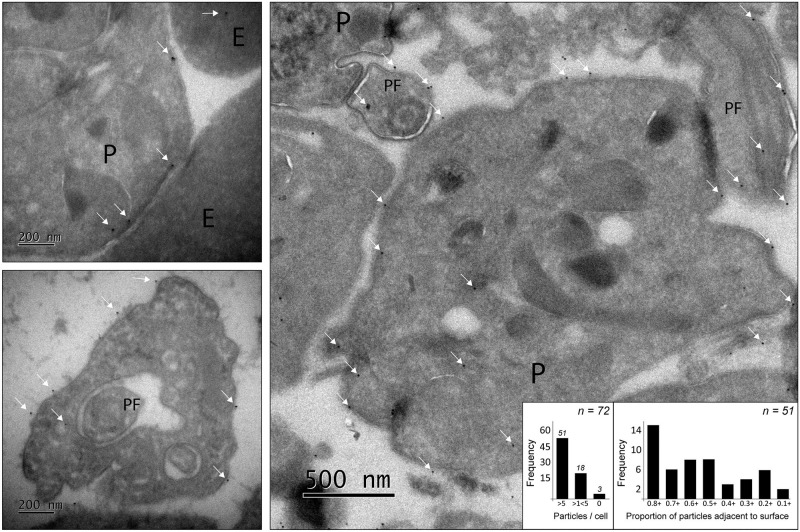
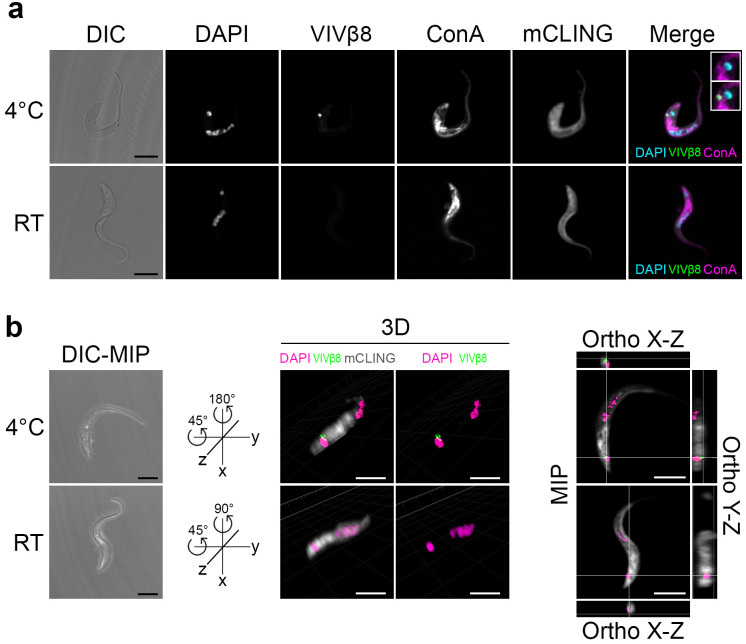
Trypanosoma vivax is a unicellular hemoparasite, and a principal cause of animal African trypanosomiasis (AAT), a vector-borne and potentially fatal livestock disease across sub-Saharan Africa. Previously, we identified diverse T. vivax-specific genes that were predicted to encode cell surface proteins. Here, we examine the immune responses of naturally and experimentally infected hosts to these unique parasite antigens, to identify immunogens that could become vaccine candidates. Immunoprofiling of host serum shows that one particular family (Fam34) elicits a consistent IgG antibody response. This gene family, which we now call Vivaxin, encodes at least 124 transmembrane glycoproteins that display quite distinct expression profiles and patterns of genetic variation. We focused on one gene (viv-β8) that encodes one particularly immunogenic vivaxin protein and which is highly expressed during infections but displays minimal polymorphism across the parasite population. Vaccination of mice with VIVβ8 adjuvanted with Quil-A elicits a strong, balanced immune response and delays parasite proliferation in some animals but, ultimately, it does not prevent disease. Although VIVβ8 is localized across the cell body and flagellar membrane, live immunostaining indicates that VIVβ8 is largely inaccessible to antibody in vivo. However, our phylogenetic analysis shows that vivaxin includes other antigens shown recently to induce immunity against T. vivax. Thus, the introduction of vivaxin represents an important advance in our understanding of the T. vivax cell surface. Besides being a source of proven and promising vaccine antigens, the gene family is clearly an important component of the parasite glycocalyx, with potential to influence host-parasite interactions.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: DA and GJW are named inventors on patents relating to the use of IFX and V31 as animal trypanosomiasis vaccines.
Figures
Similar articles
-
Variant antigen diversity in Trypanosoma vivax is not driven by recombination.Nat Commun. 2020 Feb 12;11(1):844. doi: 10.1038/s41467-020-14575-8. Nat Commun. 2020. PMID: 32051413 Free PMC article.
-
Antigenic diversity is generated by distinct evolutionary mechanisms in African trypanosome species.Proc Natl Acad Sci U S A. 2012 Feb 28;109(9):3416-21. doi: 10.1073/pnas.1117313109. Epub 2012 Feb 13. Proc Natl Acad Sci U S A. 2012. PMID: 22331916 Free PMC article.
-
Host parasite relationships in Trypanosoma (duttonella) vivax with special reference to the influence of antigenic variation.Vet Q. 1982 Jan;4(1):32-5. doi: 10.1080/01652176.1982.9693835. Vet Q. 1982. PMID: 15861585 Review.
-
Variant surface glycoproteins from Venezuelan trypanosome isolates are recognized by sera from animals infected with either Trypanosoma evansi or Trypanosoma vivax.Vet Parasitol. 2015 Jan 15;207(1-2):17-33. doi: 10.1016/j.vetpar.2014.11.004. Epub 2014 Nov 13. Vet Parasitol. 2015. PMID: 25468674 Free PMC article.
-
A Host-Pathogen Interaction Reduced to First Principles: Antigenic Variation in T. brucei.Results Probl Cell Differ. 2015;57:23-46. doi: 10.1007/978-3-319-20819-0_2. Results Probl Cell Differ. 2015. PMID: 26537376 Review.
Cited by
-
Trypanosoma vivax elicits both Th1 and Th2 immunological responses in experimentally infected cattle.PLoS One. 2025 Jul 31;20(7):e0329459. doi: 10.1371/journal.pone.0329459. eCollection 2025. PLoS One. 2025. PMID: 40743218 Free PMC article.
-
Animal Trypanosomiasis: Challenges and Prospects for New Vaccination Strategies.Microorganisms. 2024 Dec 13;12(12):2575. doi: 10.3390/microorganisms12122575. Microorganisms. 2024. PMID: 39770779 Free PMC article. Review.
References
-
- Gardiner PR. Recent studies of the biology of Trypanosoma vivax. Adv Parasitol. 1989; 28:229–317. - PubMed
-
- Osório ALAR, Madruga CR, Desquesnes M, Soares CO, Ribeiro LRR, da Costa SCG. Trypanosoma (Duttonella) vivax: Its biology, epidemiology, pathogenesis, and introduction in the New World—A review. Mem. Inst. Oswaldo Cruz. 2008; 103:1–13. - PubMed
-
- Batista JS, Rodrigues CM, Olinda RG, Silva TM, Vale RG, Câmara AC, et al.. Highly debilitating natural Trypanosoma vivax infections in Brazilian calves: epidemiology, pathology, and probable transplacental transmission. Parasitol Res. 2012; 110(1):73–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
