

Microglial control of neuronal development via somatic purinergic junctions
- PMID: 36130488
- PMCID: PMC9513806
- DOI: 10.1016/j.celrep.2022.111369
Microglial control of neuronal development via somatic purinergic junctions
Abstract
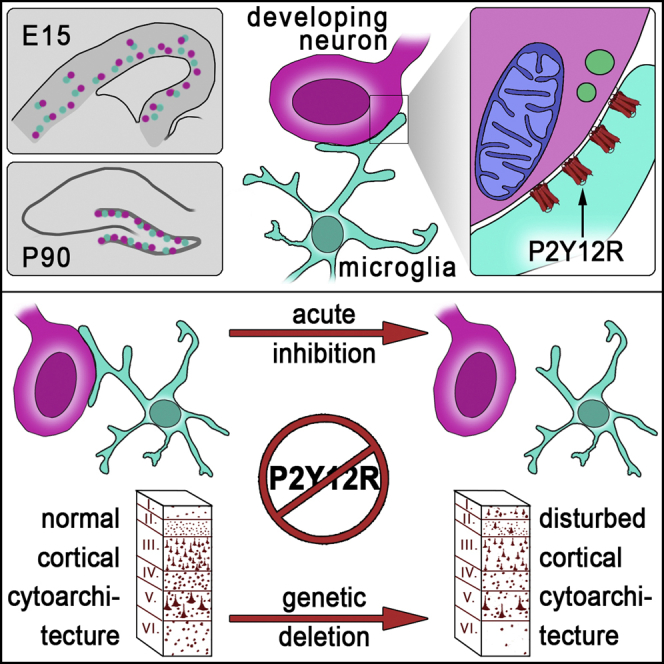
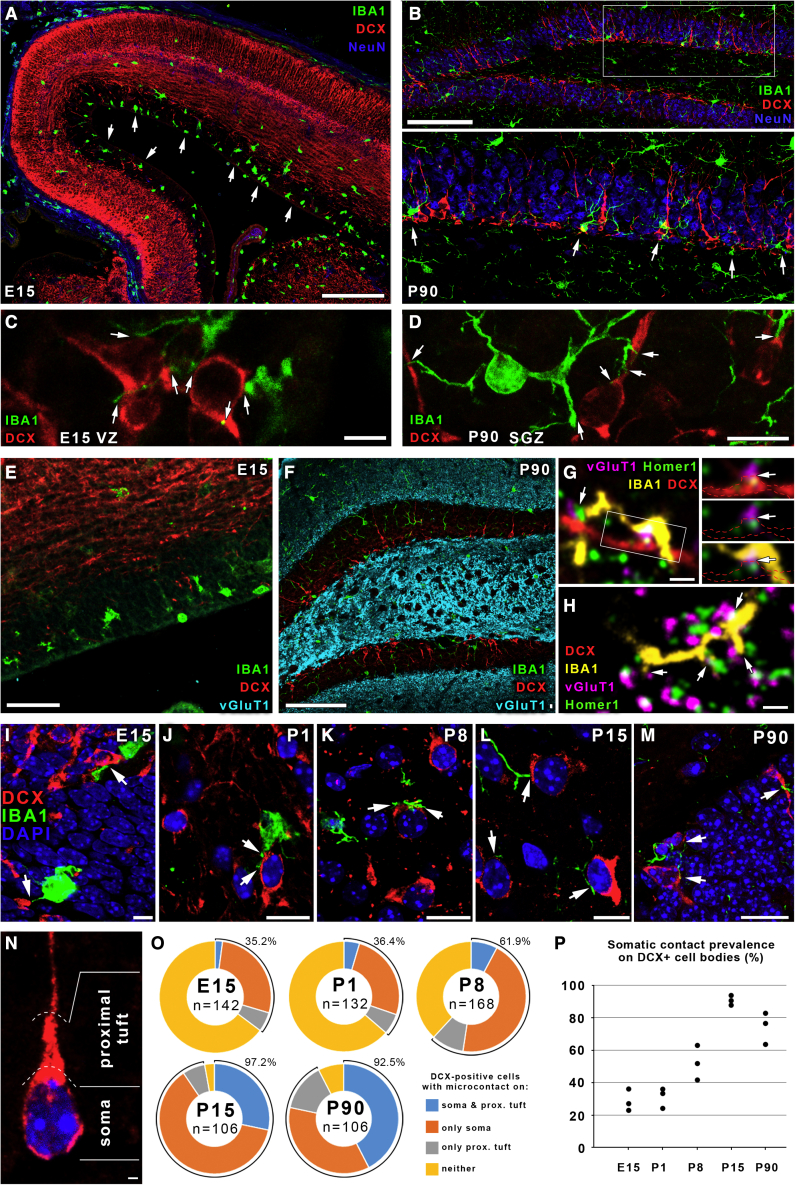
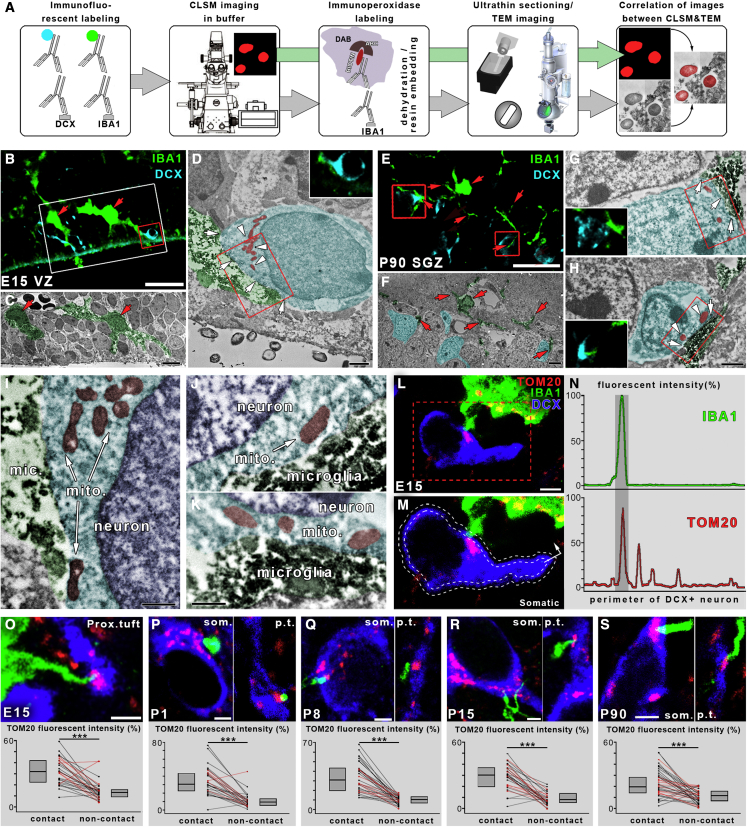
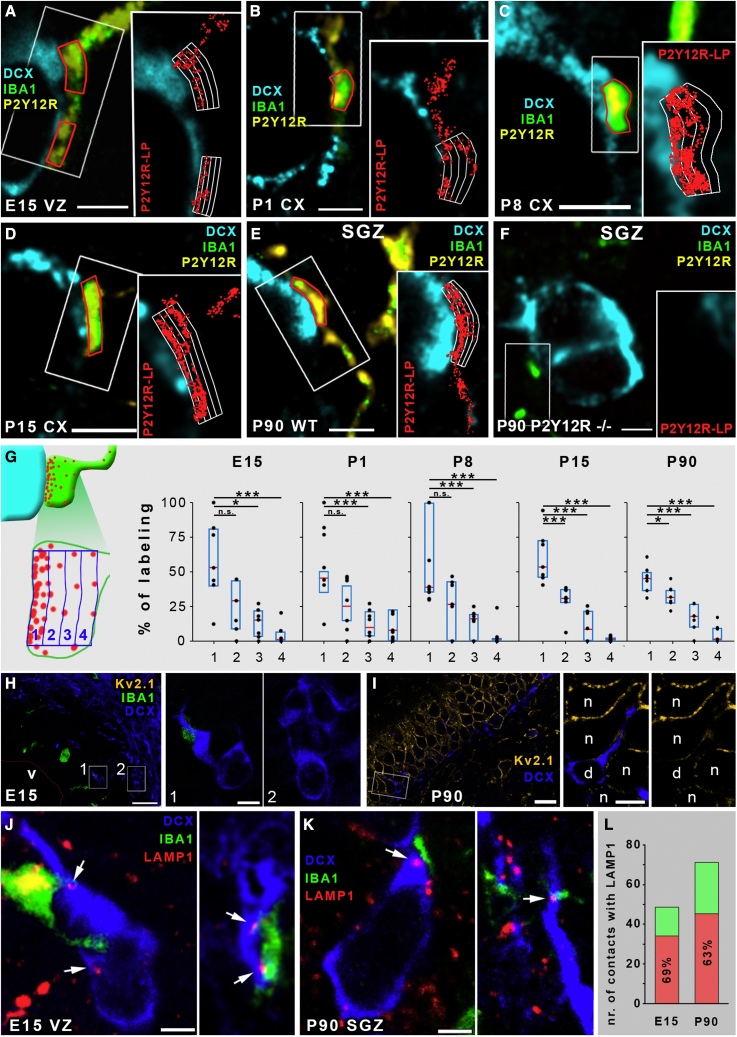
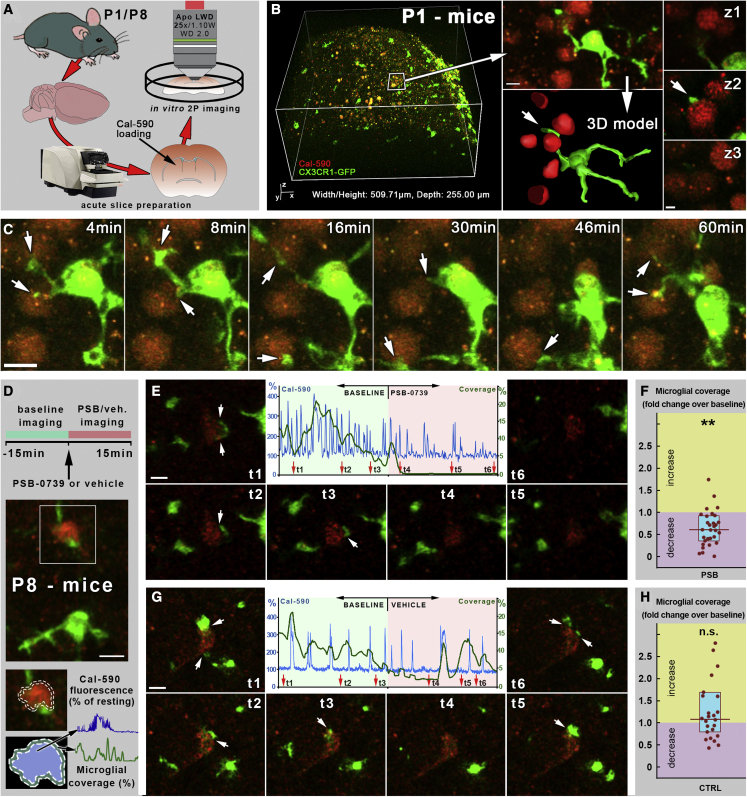
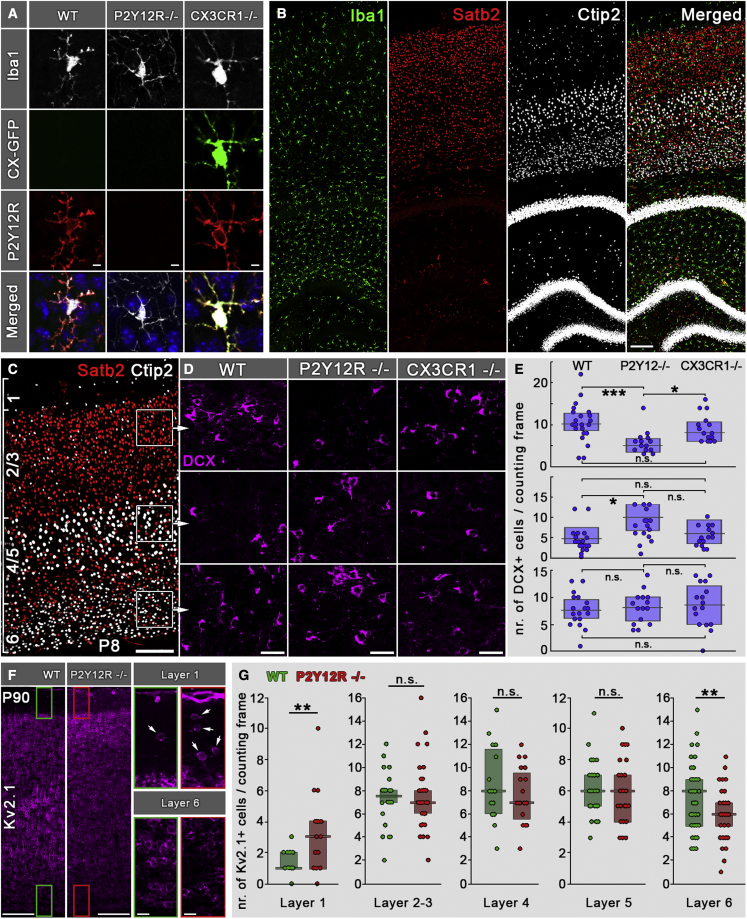
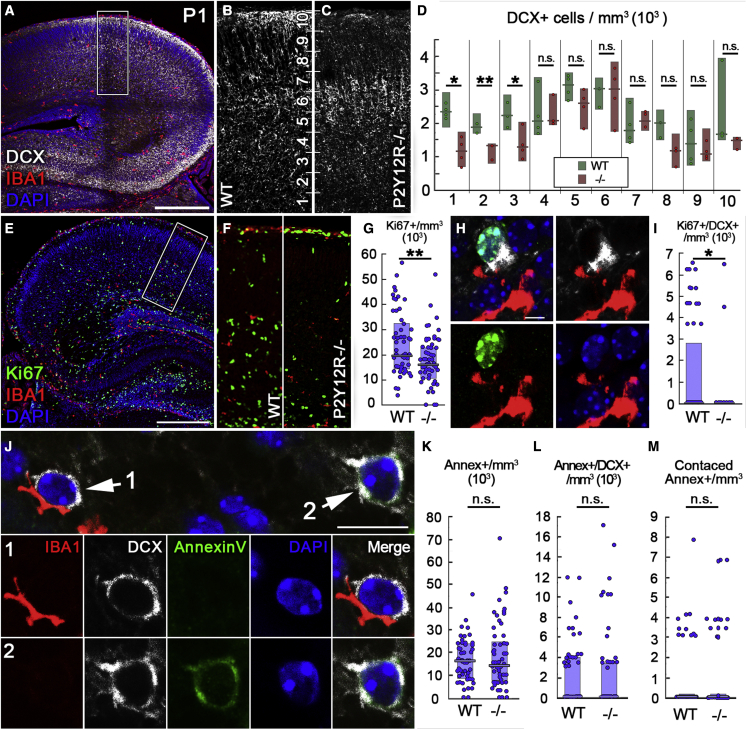
Microglia, the resident immune cells of the brain, play important roles during development. Although bi-directional communication between microglia and neuronal progenitors or immature neurons has been demonstrated, the main sites of interaction and the underlying mechanisms remain elusive. By using advanced methods, here we provide evidence that microglial processes form specialized contacts with the cell bodies of developing neurons throughout embryonic, early postnatal, and adult neurogenesis. These early developmental contacts are highly reminiscent of somatic purinergic junctions that are instrumental for microglia-neuron communication in the adult brain. The formation and maintenance of these junctions is regulated by functional microglial P2Y12 receptors, and deletion of P2Y12Rs disturbs proliferation of neuronal precursors and leads to aberrant cortical cytoarchitecture during development and in adulthood. We propose that early developmental formation of somatic purinergic junctions represents an important interface for microglia to monitor the status of immature neurons and control neurodevelopment.
Keywords: CP: Neuroscience; P2Y12R; doublecortin; microglia; neuronal development; purinergic; somatic junction.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Microglia monitor and protect neuronal function through specialized somatic purinergic junctions.Science. 2020 Jan 31;367(6477):528-537. doi: 10.1126/science.aax6752. Epub 2019 Dec 12. Science. 2020. PMID: 31831638
-
Microglial P2Y12 Receptor Regulates Seizure-Induced Neurogenesis and Immature Neuronal Projections.J Neurosci. 2019 Nov 20;39(47):9453-9464. doi: 10.1523/JNEUROSCI.0487-19.2019. Epub 2019 Oct 9. J Neurosci. 2019. PMID: 31597724 Free PMC article.
-
Mechanisms underlying microglial colonization of developing neural retina in zebrafish.Elife. 2021 Dec 7;10:e70550. doi: 10.7554/eLife.70550. Elife. 2021. PMID: 34872632 Free PMC article.
-
The behavior and functions of embryonic microglia.Anat Sci Int. 2022 Jan;97(1):1-14. doi: 10.1007/s12565-021-00631-w. Epub 2021 Sep 19. Anat Sci Int. 2022. PMID: 34537900 Free PMC article. Review.
-
Bidirectional microglia-neuron communication in the healthy brain.Neural Plast. 2013;2013:456857. doi: 10.1155/2013/456857. Epub 2013 Sep 2. Neural Plast. 2013. PMID: 24078884 Free PMC article. Review.
Cited by
-
Diagnostic and therapeutic value of P2Y12R in epilepsy.Front Pharmacol. 2023 May 10;14:1179028. doi: 10.3389/fphar.2023.1179028. eCollection 2023. Front Pharmacol. 2023. PMID: 37234715 Free PMC article. Review.
-
Microglia maintain structural integrity during fetal brain morphogenesis.Cell. 2024 Feb 15;187(4):962-980.e19. doi: 10.1016/j.cell.2024.01.012. Epub 2024 Feb 2. Cell. 2024. PMID: 38309258 Free PMC article.
-
The Signaling Pathway of the ADP Receptor P2Y12 in the Immune System: Recent Discoveries and New Challenges.Int J Mol Sci. 2023 Apr 4;24(7):6709. doi: 10.3390/ijms24076709. Int J Mol Sci. 2023. PMID: 37047682 Free PMC article. Review.
-
Microglia contribute to neuronal synchrony despite endogenous ATP-related phenotypic transformation in acute mouse brain slices.Nat Commun. 2024 Jun 26;15(1):5402. doi: 10.1038/s41467-024-49773-1. Nat Commun. 2024. PMID: 38926390 Free PMC article.
-
Microglia and Astrocytes in Postnatal Neural Circuit Formation.Glia. 2025 Feb;73(2):232-250. doi: 10.1002/glia.24650. Epub 2024 Nov 21. Glia. 2025. PMID: 39568399 Free PMC article. Review.
References
-
- Ali A.A.H., Abdel-Hafiz L., Tundo-Lavalle F., Hassan S.A., von Gall C. P2Y2 deficiency impacts adult neurogenesis and related forebrain functions. FASEB J. 2021;35:e21546. - PubMed
-
- Arrázola M.S., Andraini T., Szelechowski M., Mouledous L., Arnauné-Pelloquin L., Davezac N., Belenguer P., Rampon C., Miquel M.C. Mitochondria in developmental and adult neurogenesis. Neurotox. Res. 2019;36:257–267. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
