

Proteolytic modulation of tumor microenvironment signals during cancer progression
- PMID: 36132127
- PMCID: PMC9483212
- DOI: 10.3389/fonc.2022.935231
Proteolytic modulation of tumor microenvironment signals during cancer progression
Abstract
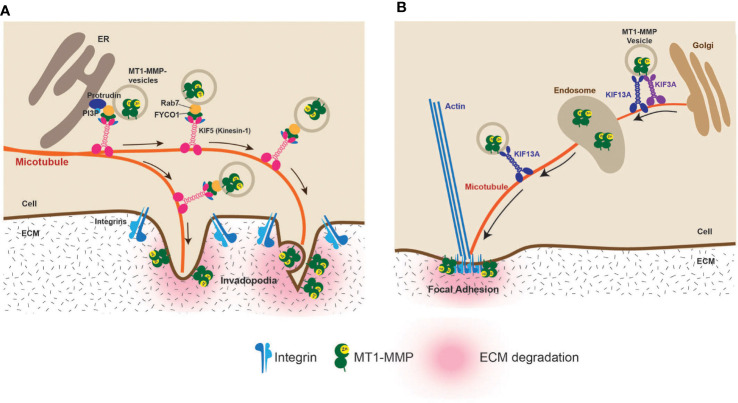
Under normal conditions, the cellular microenvironment is optimized for the proper functioning of the tissues and organs. Cells recognize and communicate with the surrounding cells and extracellular matrix to maintain homeostasis. When cancer arises, the cellular microenvironment is modified to optimize its malignant growth, evading the host immune system and finding ways to invade and metastasize to other organs. One means is a proteolytic modification of the microenvironment and the signaling molecules. It is now well accepted that cancer progression relies on not only the performance of cancer cells but also the surrounding microenvironment. This mini-review discusses the current understanding of the proteolytic modification of the microenvironment signals during cancer progression.
Keywords: ECM; Soluble factors; TME; invasion; matrikine; membrane protein shedding; proteinases.
Copyright © 2022 Itoh.
Conflict of interest statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



References
Publication types
LinkOut - more resources
Full Text Sources