

Renal oncometabolite L-2-hydroxyglutarate imposes a block in kidney tubulogenesis: Evidence for an epigenetic basis for the L-2HG-induced impairment of differentiation
- PMID: 36133305
- PMCID: PMC9483015
- DOI: 10.3389/fendo.2022.932286
Renal oncometabolite L-2-hydroxyglutarate imposes a block in kidney tubulogenesis: Evidence for an epigenetic basis for the L-2HG-induced impairment of differentiation
Abstract
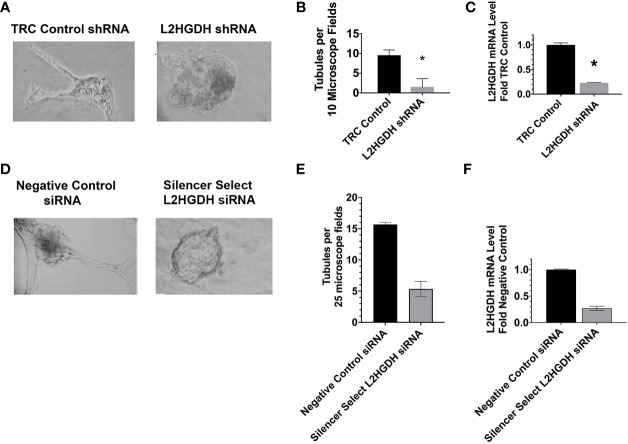
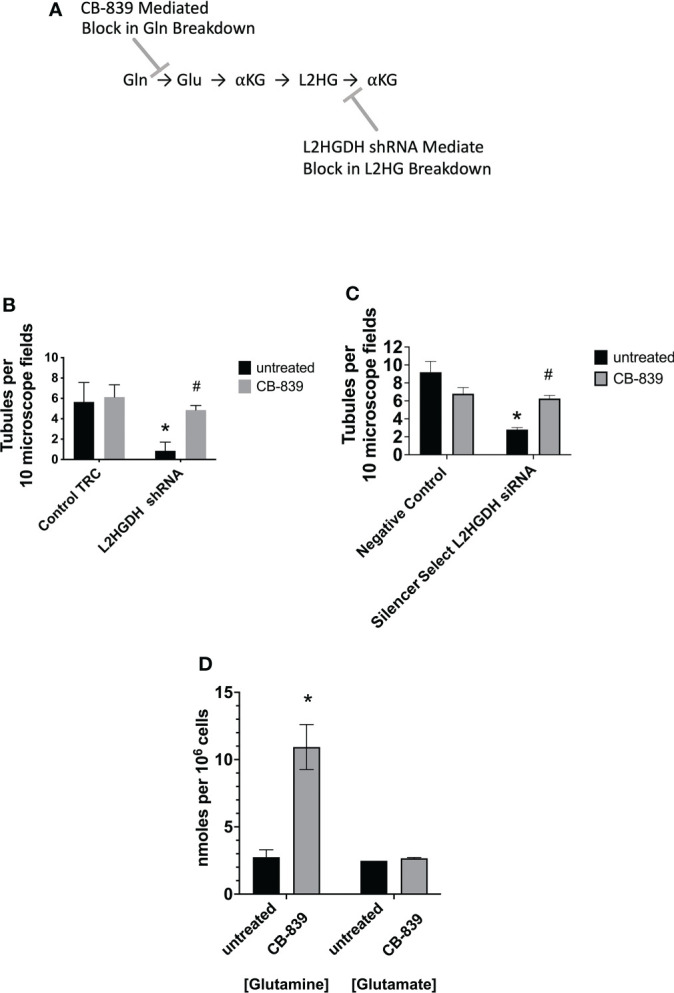
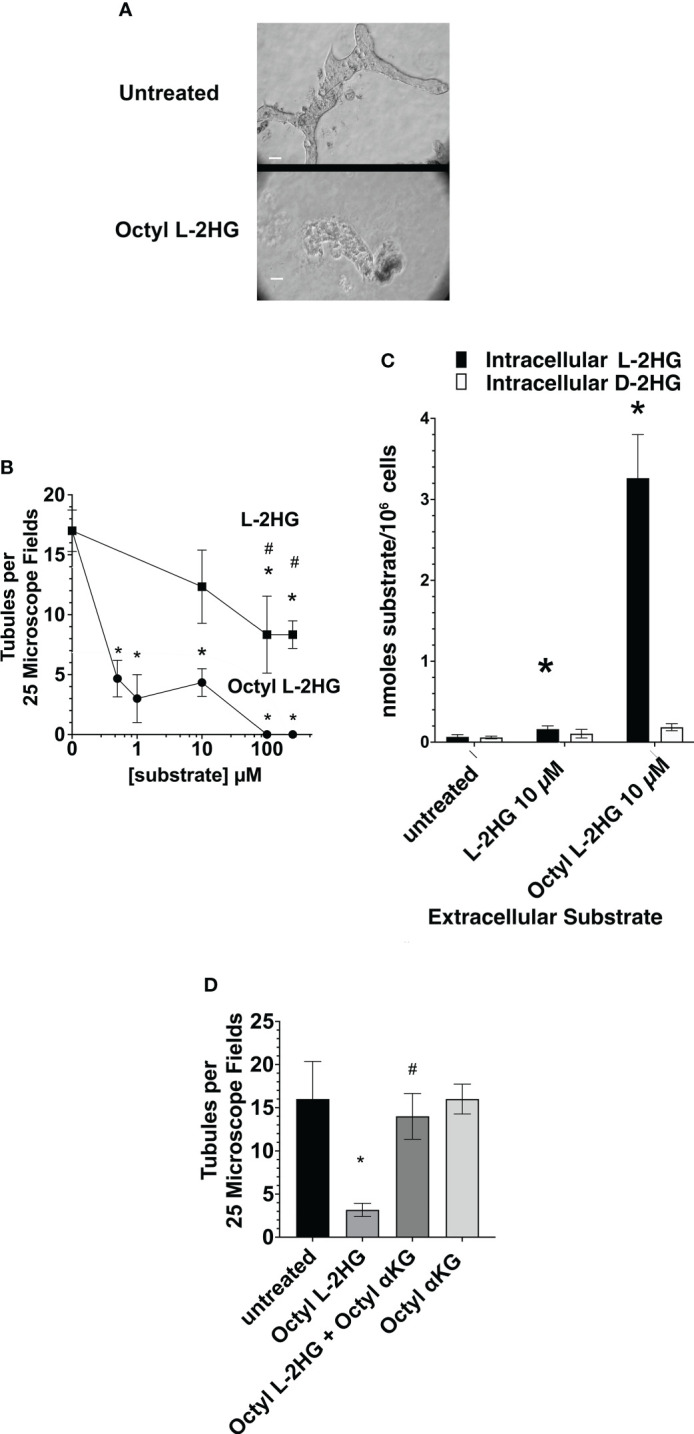
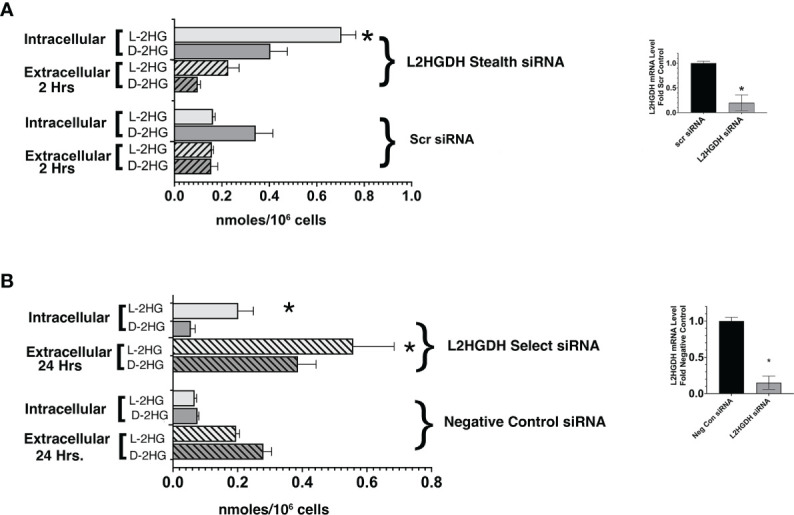
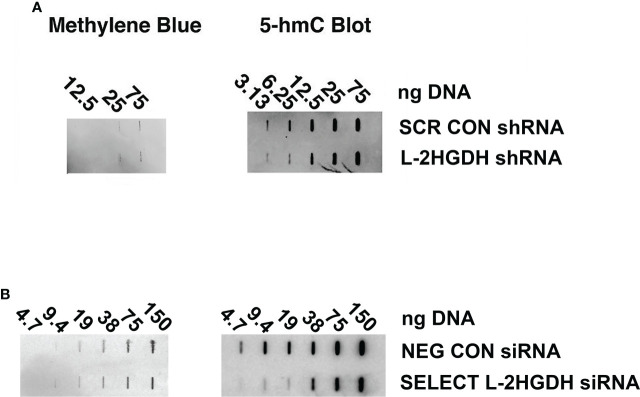
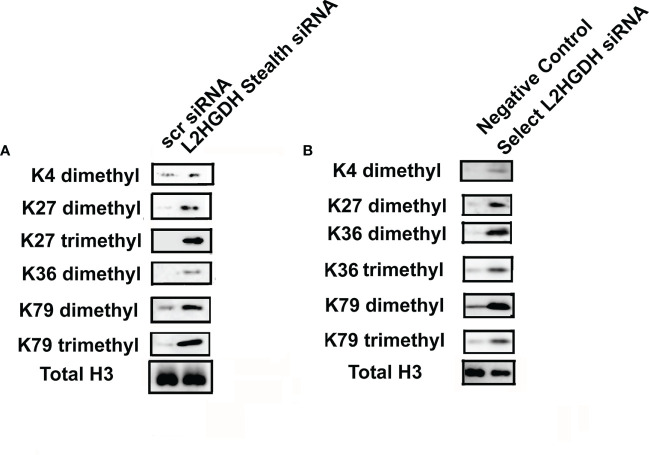
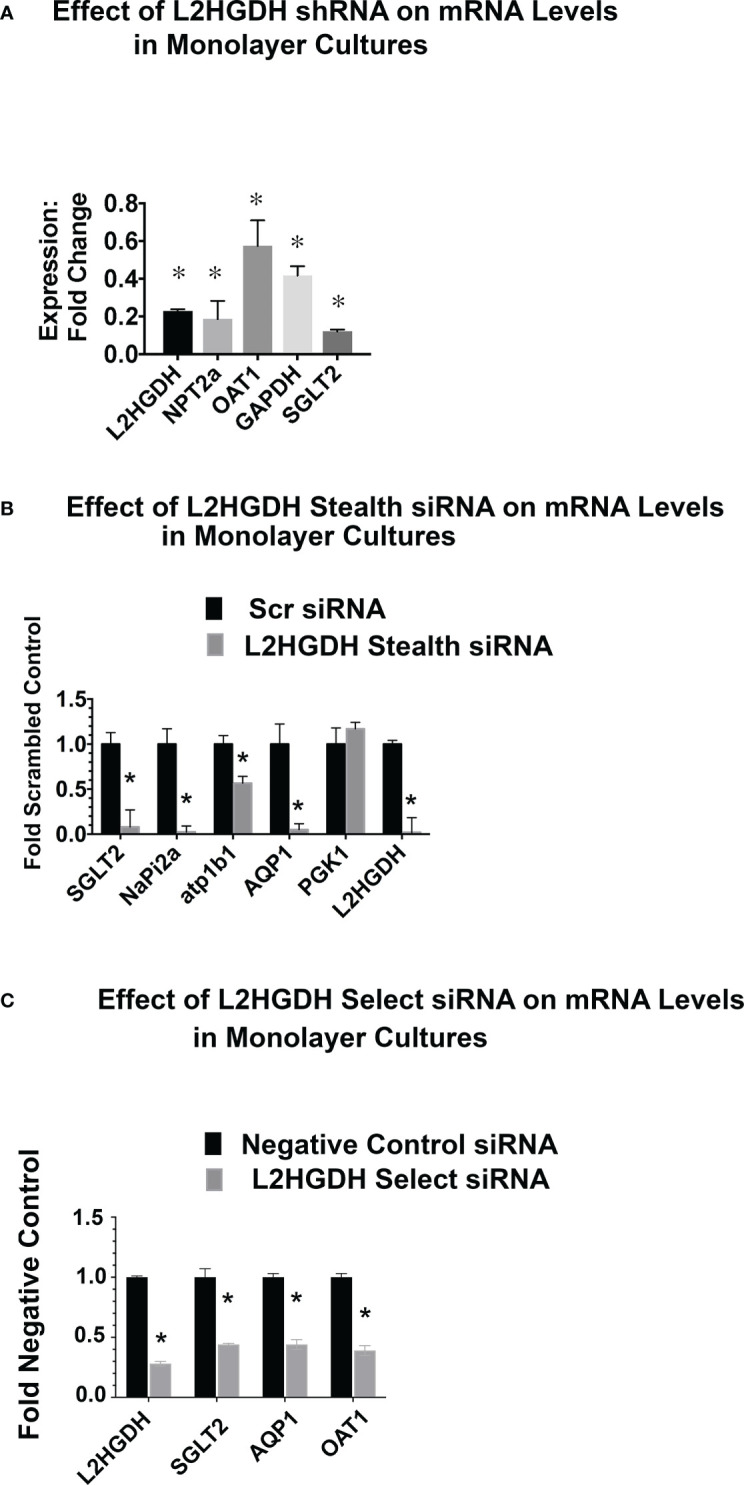
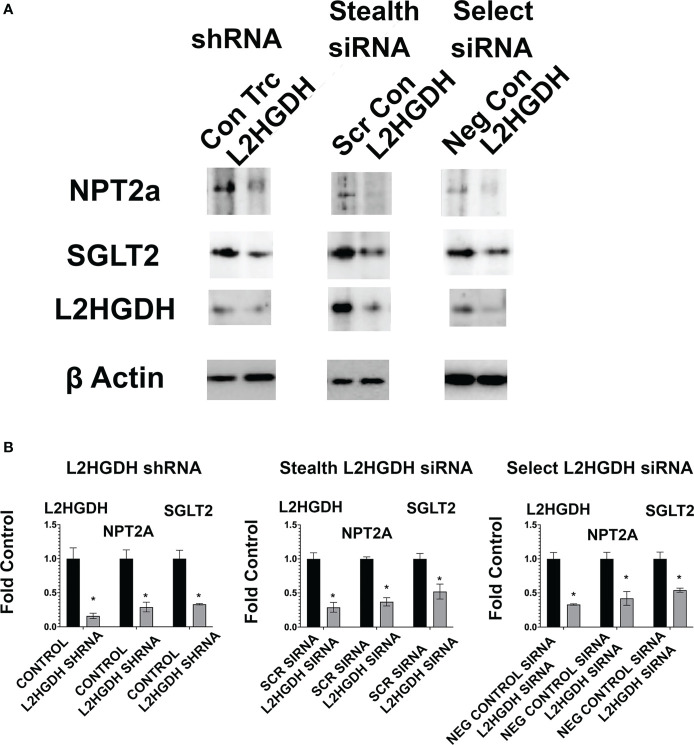
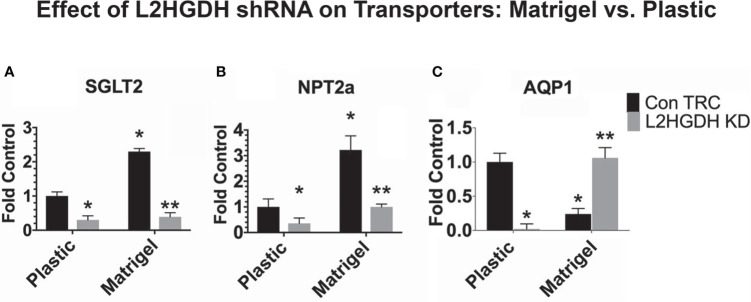
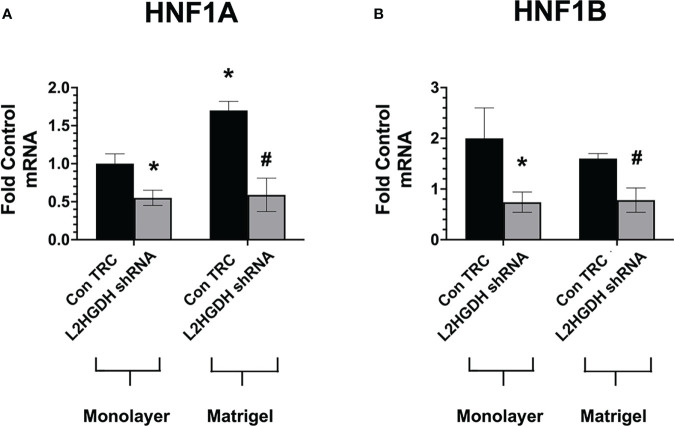
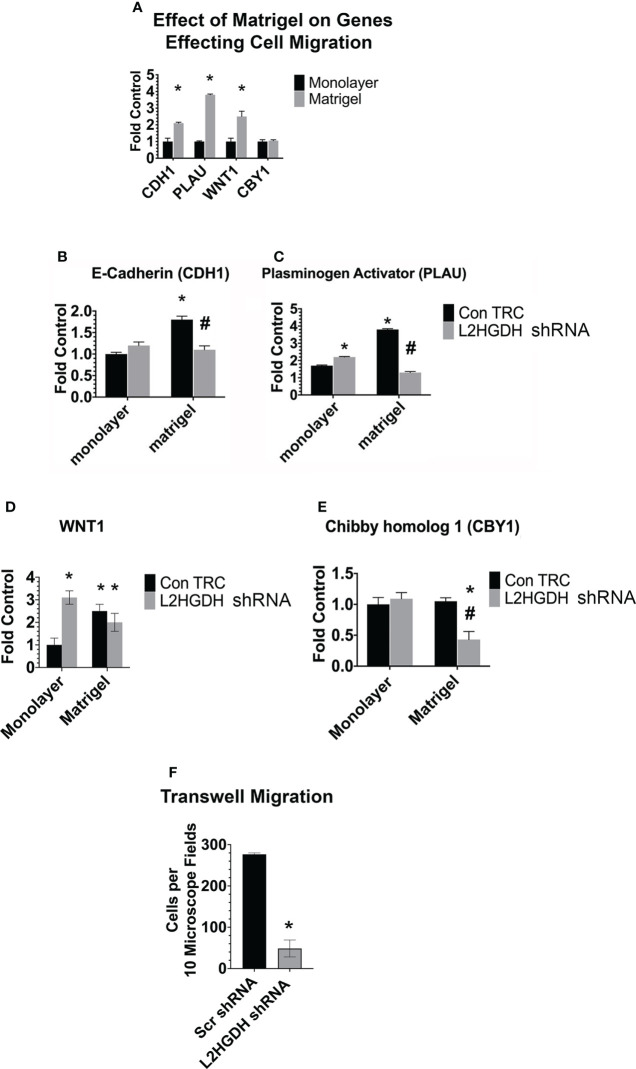
2-Hydroxyglutarate (2HG) overproducing tumors arise in a number of tissues, including the kidney. The tumorigenesis resulting from overproduced 2HG has been attributed to the ability of 2HG alter gene expression by inhibiting α-ketoglutarate (αKG)-dependent dioxygenases, including Ten-eleven-Translocation (TET) enzymes. Genes that regulate cellular differentiation are reportedly repressed, blocking differentiation of mesenchymal cells into myocytes, and adipocytes. In this report, the expression of the enzyme responsible for L2HG degradation, L-2HG dehydrogenase (L2HGDH), is knocked down, using lentiviral shRNA, as well as siRNA, in primary cultures of normal Renal Proximal Tubule (RPT) cells. The knockdown (KD) results in increased L-2HG levels, decreased demethylation of 5mC in genomic DNA, and increased methylation of H3 Histones. Consequences include reduced tubulogenesis by RPT cells in matrigel, and reduced expression of molecular markers of differentiation, including membrane transporters as well as HNF1α and HNF1β, which regulate their transcription. These results are consistent with the hypothesis that oncometabolite 2HG blocks RPT differentiation by altering the methylation status of chromatin in a manner that impedes the transcriptional events required for normal differentiation. Presumably, similar alterations are responsible for promoting the expansion of renal cancer stem-cells, increasing their propensity for malignant transformation.
Keywords: L-2-Hydroxyglutarate; differentiation; matrigel (MA); proximal tubule; renal cell carcinoma.
Copyright © 2022 Taub, Mahmoudzadeh, Tennessen and Sudarshan.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources