

Global drivers of variation in cup nest size in passerine birds
- PMID: 36134498
- PMCID: PMC10092846
- DOI: 10.1111/1365-2656.13815
Global drivers of variation in cup nest size in passerine birds
Abstract
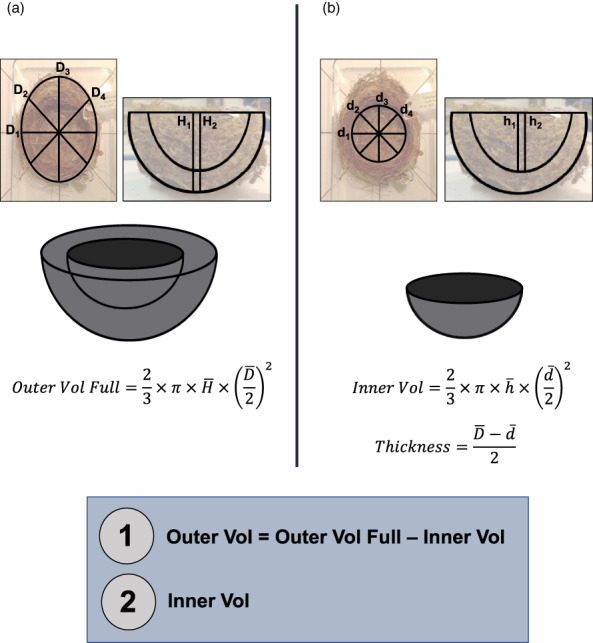
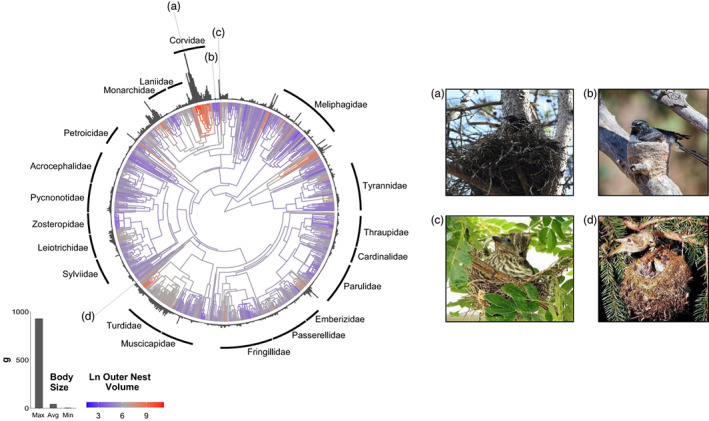
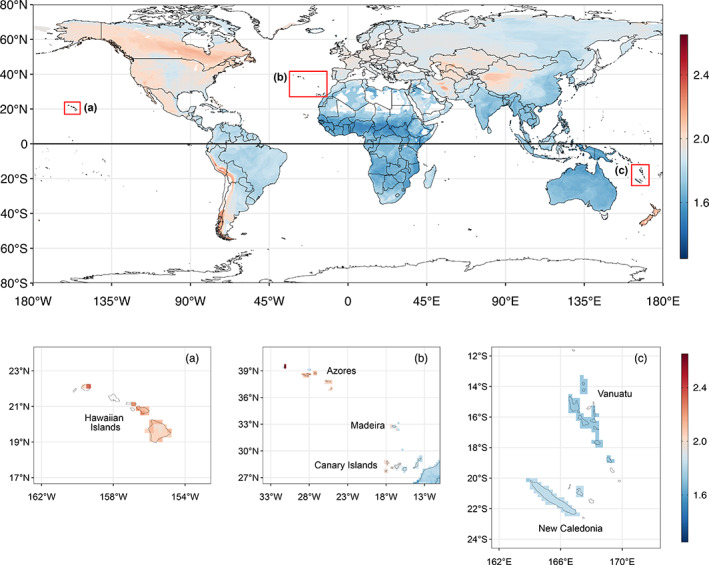
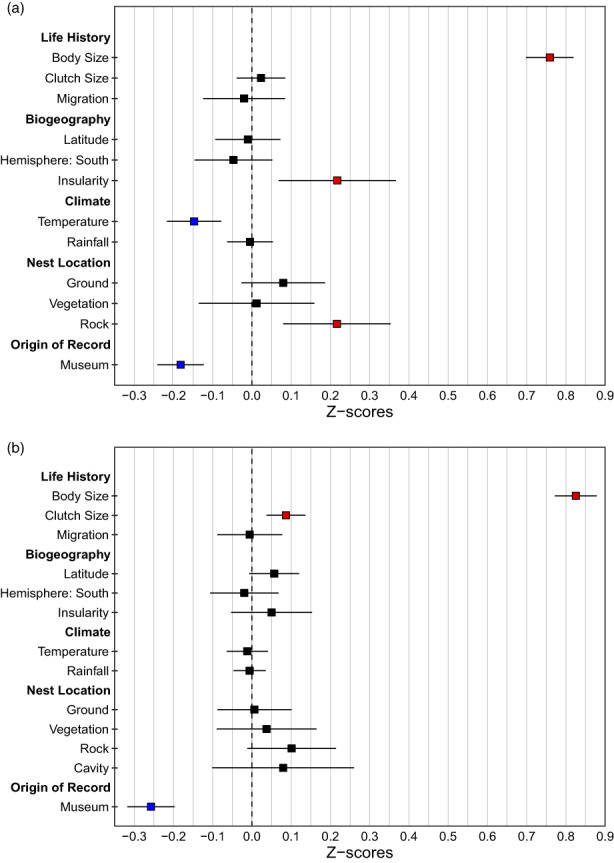
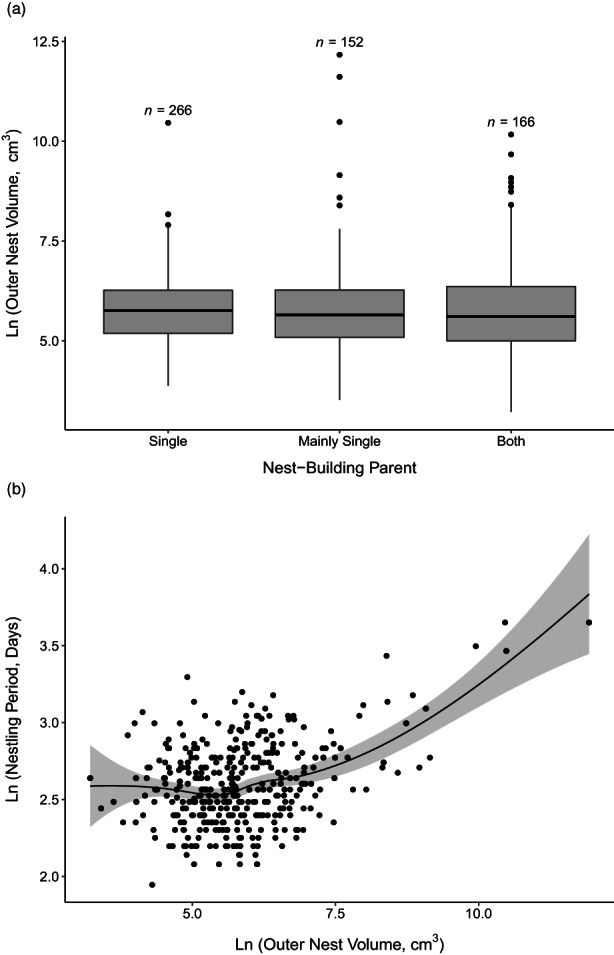
The size of a bird's nest can play a key role in ensuring reproductive success and is determined by a variety of factors. The primary function of the nest is to protect offspring from the environment and predators. Field studies in a number of passerine species have indicated that higher-latitude populations in colder habitats build larger nests with thicker walls compared to lower-latitude populations, but that these larger nests are more vulnerable to predation. Increases in nest size can also be driven by sexual selection, as nest size can act as a signal of parental quality and prompt differential investment in other aspects of care. It is unknown, however, how these microevolutionary patterns translate to a macroevolutionary scale. Here, we investigate potential drivers of variation in the outer and inner volume of open cup nests using a large dataset of nest measurements from 1117 species of passerines breeding in a diverse range of environments. Our dataset is sourced primarily from the nest specimens at the Natural History Museum (UK), complemented with information from ornithological handbooks and online databases. We use phylogenetic comparative methods to test long-standing hypotheses about potential macroevolutionary correlates of nest size, namely nest location, clutch size and variables relating to parental care, together with environmental and geographical factors such as temperature, rainfall, latitude and insularity. After controlling for phylogeny and parental body size, we demonstrate that the outer volume of the nest is greater in colder climates, in island-dwelling species and in species that nest on cliffs or rocks. By contrast, the inner cup volume is associated solely with average clutch size, increasing with the number of chicks raised in the nest. We do not find evidence that nest size is related to the length of parental care for nestlings. Our study reveals that the average temperature in the breeding range, along with several key life-history traits and proxies of predation threat, shapes the global interspecific variation in passerine cup nest size. We also showcase the utility of museum nest collections-a historically underused resource-for large-scale studies of trait evolution.
Keywords: differential allocation hypothesis; museum collections; nest size; parental investment; passerine nests; predation threat.
© 2022 The Authors. Journal of Animal Ecology published by John Wiley & Sons Ltd on behalf of British Ecological Society.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Evolution of nest architecture in tyrant flycatchers and allies.Philos Trans R Soc Lond B Biol Sci. 2023 Aug 28;378(1884):20220148. doi: 10.1098/rstb.2022.0148. Epub 2023 Jul 10. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37427478 Free PMC article.
-
Elevation and latitude interact to drive life-history variation in precocial birds: a comparative analysis using galliformes.J Anim Ecol. 2016 Nov;85(6):1528-1539. doi: 10.1111/1365-2656.12570. Epub 2016 Aug 18. J Anim Ecol. 2016. PMID: 27392151
-
Development syndromes in New World temperate and tropical songbirds.PLoS One. 2020 Aug 17;15(8):e0233627. doi: 10.1371/journal.pone.0233627. eCollection 2020. PLoS One. 2020. PMID: 32804928 Free PMC article.
-
Predators and the breeding bird: behavioral and reproductive flexibility under the risk of predation.Biol Rev Camb Philos Soc. 2009 Aug;84(3):485-513. doi: 10.1111/j.1469-185X.2009.00085.x. Biol Rev Camb Philos Soc. 2009. PMID: 19659887 Review.
-
The Evolution of Using Shed Snake Skin in Bird Nests.Am Nat. 2025 Feb;205(2):170-183. doi: 10.1086/733208. Epub 2024 Dec 18. Am Nat. 2025. PMID: 39913940 Review.
Cited by
-
A global database of bird nest traits.Sci Data. 2023 Dec 21;10(1):923. doi: 10.1038/s41597-023-02837-1. Sci Data. 2023. PMID: 38129417 Free PMC article.
-
Beak shape and nest material use in birds.Philos Trans R Soc Lond B Biol Sci. 2023 Aug 28;378(1884):20220147. doi: 10.1098/rstb.2022.0147. Epub 2023 Jul 10. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37427471 Free PMC article.
-
The evolutionary ecology of nests: a cross-taxon approach.Philos Trans R Soc Lond B Biol Sci. 2023 Aug 28;378(1884):20220136. doi: 10.1098/rstb.2022.0136. Epub 2023 Jul 10. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37427472 Free PMC article.
-
Variation in nest-building behaviour in birds: a multi-species approach.Philos Trans R Soc Lond B Biol Sci. 2023 Aug 28;378(1884):20220145. doi: 10.1098/rstb.2022.0145. Epub 2023 Jul 10. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37427476 Free PMC article.
-
Evolution of nest architecture in tyrant flycatchers and allies.Philos Trans R Soc Lond B Biol Sci. 2023 Aug 28;378(1884):20220148. doi: 10.1098/rstb.2022.0148. Epub 2023 Jul 10. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37427478 Free PMC article.
References
-
- Antonov, A. (2004). Smaller Eastern Olivaceous Warbler Hippolais pallida elaeica nests suffer less predation than larger ones. Acta Ornithologica, 39(2), 87–92. 10.3161/068.039.0205 - DOI
-
- Barber, I. (2013). The evolutionary ecology of nest construction: Insight from recent fish studies. Avian Biology Research, 6(2), 83–98. 10.3184/175815513X13609538379947 - DOI
-
- Biddle, L. E. , Dickinson, A. M. , Broughton, R. E. , Gray, L. A. , Bennett, S. L. , Goodman, A. M. , & Deeming, D. C. (2019). Construction materials affect the hydrological properties of bird nests. Journal of Zoology, 309(3), 161–171. 10.1111/JZO.12713 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
