

Massive annotation of bacterial L-asparaginases reveals their puzzling distribution and frequent gene transfer events
- PMID: 36138049
- PMCID: PMC9500103
- DOI: 10.1038/s41598-022-19689-1
Massive annotation of bacterial L-asparaginases reveals their puzzling distribution and frequent gene transfer events
Abstract
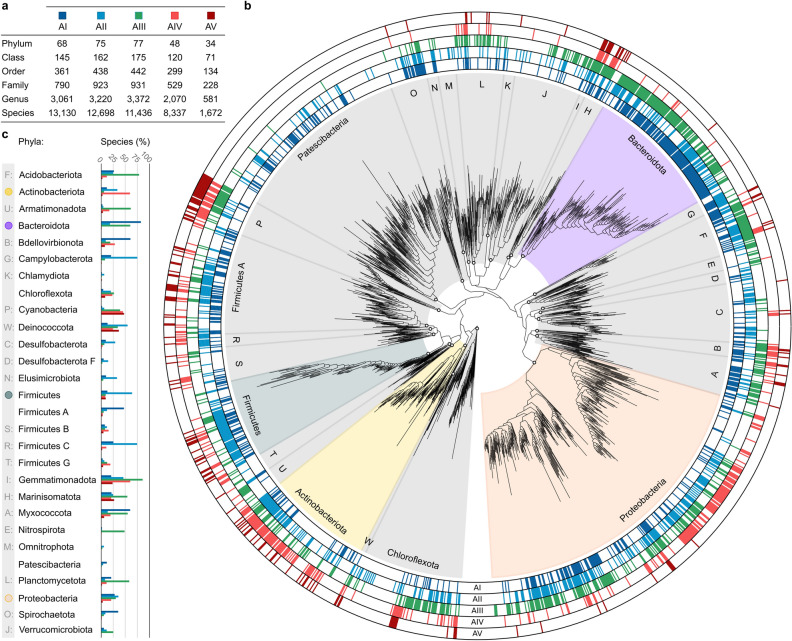
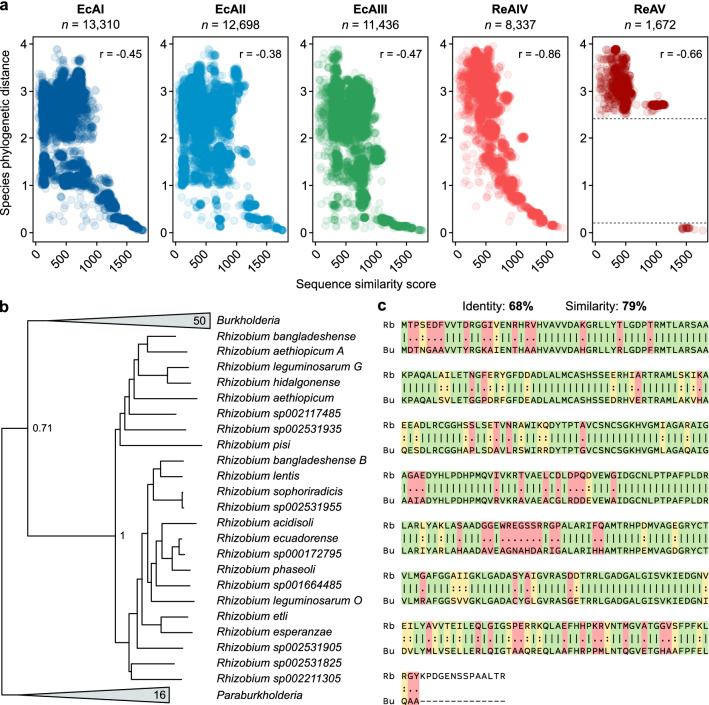
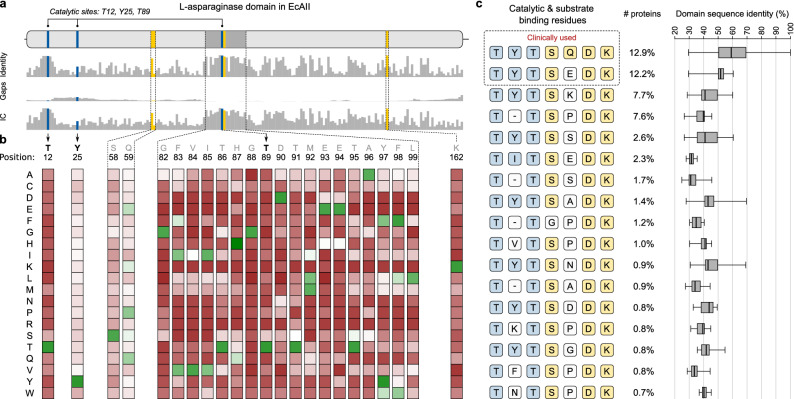
L-Asparaginases, which convert L-asparagine to L-aspartate and ammonia, come in five types, AI-AV. Some bacterial type AII enzymes are a key element in the treatment of acute lymphoblastic leukemia in children, but new L-asparaginases with better therapeutic properties are urgently needed. Here, we search publicly available bacterial genomes to annotate L-asparaginase proteins belonging to the five known types. We characterize taxonomic, phylogenetic, and genomic patterns of L-asparaginase occurrences pointing to frequent horizontal gene transfer (HGT) events, also occurring multiple times in the same recipient species. We show that the reference AV gene, encoding a protein originally found and structurally studied in Rhizobium etli, was acquired via HGT from Burkholderia. We also describe the sequence variability of the five L-asparaginase types and map the conservation levels on the experimental or predicted structures of the reference enzymes, finding the most conserved residues in the protein core near the active site, and the most variable ones on the protein surface. Additionally, we highlight the most common sequence features of bacterial AII proteins that may aid in selecting therapeutic L-asparaginases. Finally, we point to taxonomic units of bacteria that do not contain recognizable sequences of any of the known L-asparaginase types, implying that those microorganisms most likely contain new, as yet unknown types of L-asparaginases. Such novel enzymes, when properly identified and characterized, could hold promise as antileukemic drugs.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
The L-asparagine operon of Rhizobium etli contains a gene encoding an atypical asparaginase.FEMS Microbiol Lett. 2000 Aug 15;189(2):177-82. doi: 10.1111/j.1574-6968.2000.tb09226.x. FEMS Microbiol Lett. 2000. PMID: 10930734
-
Crystal structure of active site mutant of antileukemic L-asparaginase reveals conserved zinc-binding site.FEBS J. 2014 Sep;281(18):4097-111. doi: 10.1111/febs.12906. Epub 2014 Jul 28. FEBS J. 2014. PMID: 25040257
-
A critical review on properties and applications of microbial l-asparaginases.Crit Rev Microbiol. 2016 Sep;42(5):720-37. doi: 10.3109/1040841X.2015.1022505. Epub 2015 Apr 13. Crit Rev Microbiol. 2016. PMID: 25865363 Review.
-
Microbial L-asparaginase: purification, characterization and applications.Arch Microbiol. 2020 Jul;202(5):967-981. doi: 10.1007/s00203-020-01814-1. Epub 2020 Feb 12. Arch Microbiol. 2020. PMID: 32052094 Review.
-
Experimental Data in Support of a Direct Displacement Mechanism for Type I/II L-Asparaginases.J Biol Chem. 2016 Mar 4;291(10):5088-100. doi: 10.1074/jbc.M115.699884. Epub 2016 Jan 5. J Biol Chem. 2016. PMID: 26733195 Free PMC article.
Cited by
-
RrA, an enzyme from Rhodospirillum rubrum, is a prototype of a new family of short-chain L-asparaginases.Protein Sci. 2024 Apr;33(4):e4920. doi: 10.1002/pro.4920. Protein Sci. 2024. PMID: 38501449 Free PMC article.
-
Optimizing Gene Sources for L-asparaginase Production: A Comparative Review.Curr Microbiol. 2025 Jul 12;82(9):381. doi: 10.1007/s00284-025-04351-6. Curr Microbiol. 2025. PMID: 40646362 Review.
-
Engineering and Expression Strategies for Optimization of L-Asparaginase Development and Production.Int J Mol Sci. 2023 Oct 16;24(20):15220. doi: 10.3390/ijms242015220. Int J Mol Sci. 2023. PMID: 37894901 Free PMC article. Review.
-
The effects of nature-inspired amino acid substitutions on structural and biochemical properties of the E. coli L-asparaginase EcAIII.Protein Sci. 2023 Jun;32(6):e4647. doi: 10.1002/pro.4647. Protein Sci. 2023. PMID: 37095066 Free PMC article.
-
Probing the active site of Class 3 L-asparaginase by mutagenesis. I. Tinkering with the zinc coordination site of ReAV.Front Chem. 2024 Apr 4;12:1381032. doi: 10.3389/fchem.2024.1381032. eCollection 2024. Front Chem. 2024. PMID: 38638878 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
