

Deregulation of Mitochondrial Calcium Handling Due to Presenilin Loss Disrupts Redox Homeostasis and Promotes Neuronal Dysfunction
- PMID: 36139715
- PMCID: PMC9495597
- DOI: 10.3390/antiox11091642
Deregulation of Mitochondrial Calcium Handling Due to Presenilin Loss Disrupts Redox Homeostasis and Promotes Neuronal Dysfunction
Abstract
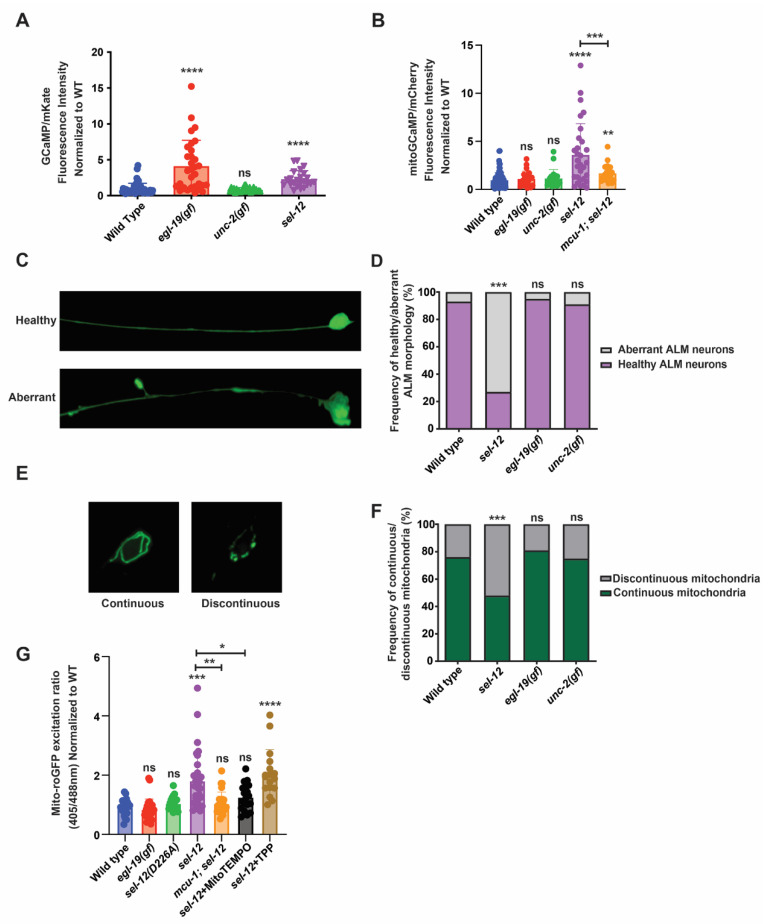
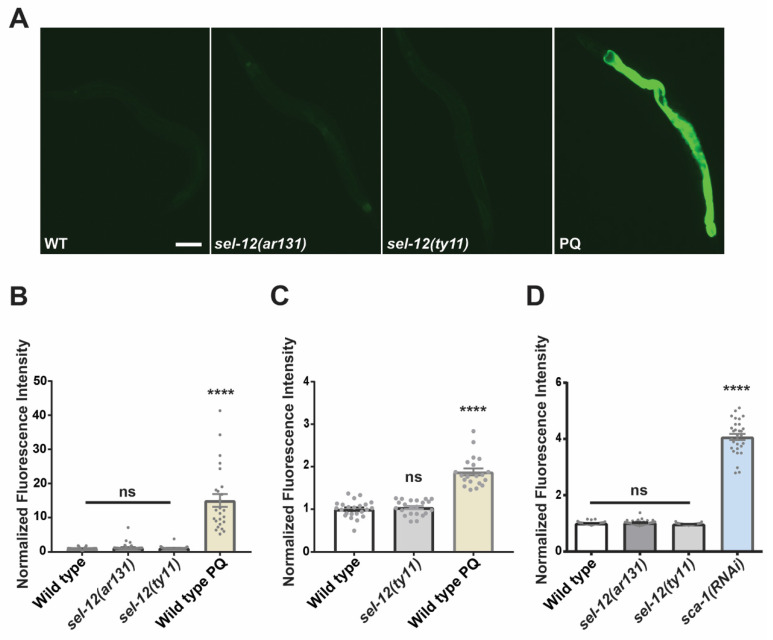
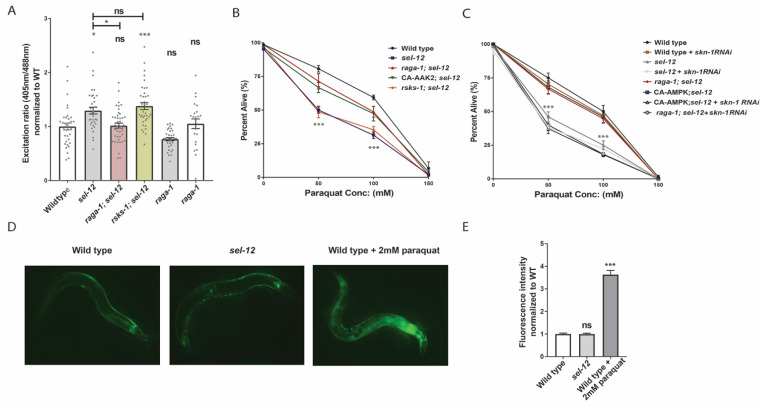
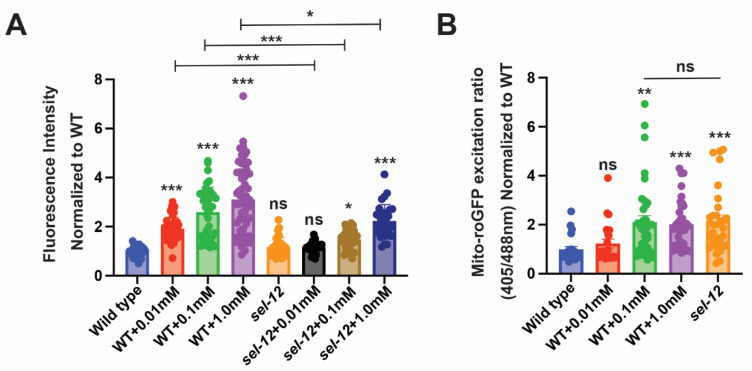
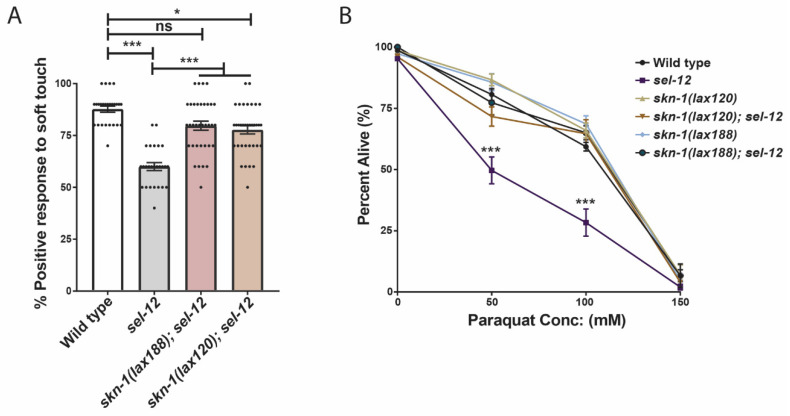
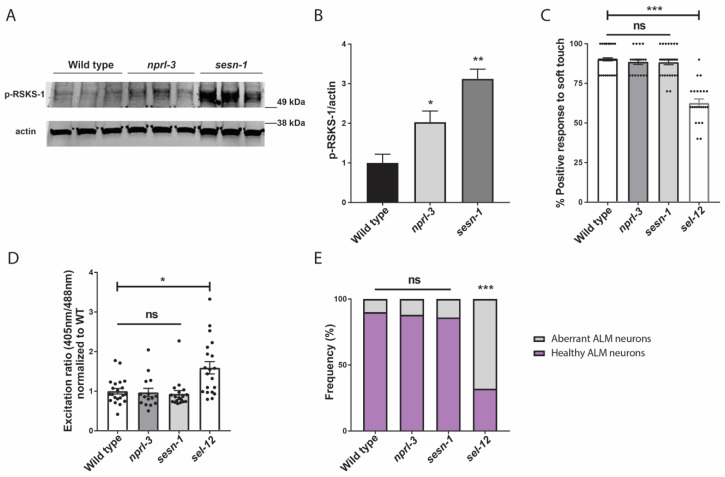
Mitochondrial dysfunction and oxidative stress are major contributors to the pathophysiology of neurodegenerative diseases, including Alzheimer's disease (AD). However, the mechanisms driving mitochondrial dysfunction and oxidative stress are unclear. Familial AD (fAD) is an early onset form of AD caused primarily by mutations in the presenilin-encoding genes. Previously, using Caenorhabditis elegans as a model system to study presenilin function, we found that loss of C. elegans presenilin orthologue SEL-12 results in elevated mitochondrial and cytosolic calcium levels. Here, we provide evidence that elevated neuronal mitochondrial generated reactive oxygen species (ROS) and subsequent neurodegeneration in sel-12 mutants are a consequence of the increase of mitochondrial calcium levels and not cytosolic calcium levels. We also identify mTORC1 signaling as a critical factor in sustaining high ROS in sel-12 mutants in part through its repression of the ROS scavenging system SKN-1/Nrf. Our study reveals that SEL-12/presenilin loss disrupts neuronal ROS homeostasis by increasing mitochondrial ROS generation and elevating mTORC1 signaling, which exacerbates this imbalance by suppressing SKN-1/Nrf antioxidant activity.
Keywords: Alzheimer’s disease; Nrf2; calcium; mitochondria; neuronal dysfunction; oxidative stress; presenilin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Increased mitochondrial calcium uptake and concomitant mitochondrial activity by presenilin loss promotes mTORC1 signaling to drive neurodegeneration.Aging Cell. 2021 Oct;20(10):e13472. doi: 10.1111/acel.13472. Epub 2021 Sep 9. Aging Cell. 2021. PMID: 34499406 Free PMC article.
-
Presenilin mutations deregulate mitochondrial Ca2+ homeostasis and metabolic activity causing neurodegeneration in Caenorhabditis elegans.Elife. 2018 Jul 10;7:e33052. doi: 10.7554/eLife.33052. Elife. 2018. PMID: 29989545 Free PMC article.
-
Corrupted ER-mitochondrial calcium homeostasis promotes the collapse of proteostasis.Aging Cell. 2020 Jan;19(1):e13065. doi: 10.1111/acel.13065. Epub 2019 Nov 12. Aging Cell. 2020. PMID: 31714672 Free PMC article.
-
Role of Presenilin in Mitochondrial Oxidative Stress and Neurodegeneration in Caenorhabditis elegans.Antioxidants (Basel). 2018 Aug 24;7(9):111. doi: 10.3390/antiox7090111. Antioxidants (Basel). 2018. PMID: 30149498 Free PMC article. Review.
-
The Role of Mitochondrial Calcium Homeostasis in Alzheimer's and Related Diseases.Int J Mol Sci. 2020 Dec 1;21(23):9153. doi: 10.3390/ijms21239153. Int J Mol Sci. 2020. PMID: 33271784 Free PMC article. Review.
Cited by
-
Identification of presenilin mutations that have sufficient gamma-secretase proteolytic activity to mediate Notch signaling but disrupt organelle and neuronal health.Neurobiol Dis. 2025 Aug;212:106961. doi: 10.1016/j.nbd.2025.106961. Epub 2025 May 20. Neurobiol Dis. 2025. PMID: 40404063 Free PMC article.
-
C. elegans Presenilin Mediates Inter-Organelle Contacts and Communication that Is Required for Lysosome Activity.Aging Dis. 2024 Feb 28;16(5):3022-3039. doi: 10.14336/AD.2024.0228. Aging Dis. 2024. PMID: 38607730 Free PMC article.
-
Modulating mitochondrial calcium channels (TRPM2/MCU/NCX) as a therapeutic strategy for neurodegenerative disorders.Front Neurosci. 2023 Oct 20;17:1202167. doi: 10.3389/fnins.2023.1202167. eCollection 2023. Front Neurosci. 2023. PMID: 37928737 Free PMC article. Review.
-
Temporal Changes in Mitochondria-Centric Excitotoxic Responses Following Severe Penetrating Traumatic Brain Injury.Biomedicines. 2025 Jun 21;13(7):1520. doi: 10.3390/biomedicines13071520. Biomedicines. 2025. PMID: 40722596 Free PMC article.
-
Subcellular Imaging of Neuronal Calcium Handling In Vivo.J Vis Exp. 2023 Mar 17;(193):10.3791/64928. doi: 10.3791/64928. J Vis Exp. 2023. PMID: 37010315 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
