

TNF-α and IL-1β Modulate Blood-Brain Barrier Permeability and Decrease Amyloid-β Peptide Efflux in a Human Blood-Brain Barrier Model
- PMID: 36142143
- PMCID: PMC9499506
- DOI: 10.3390/ijms231810235
TNF-α and IL-1β Modulate Blood-Brain Barrier Permeability and Decrease Amyloid-β Peptide Efflux in a Human Blood-Brain Barrier Model
Abstract
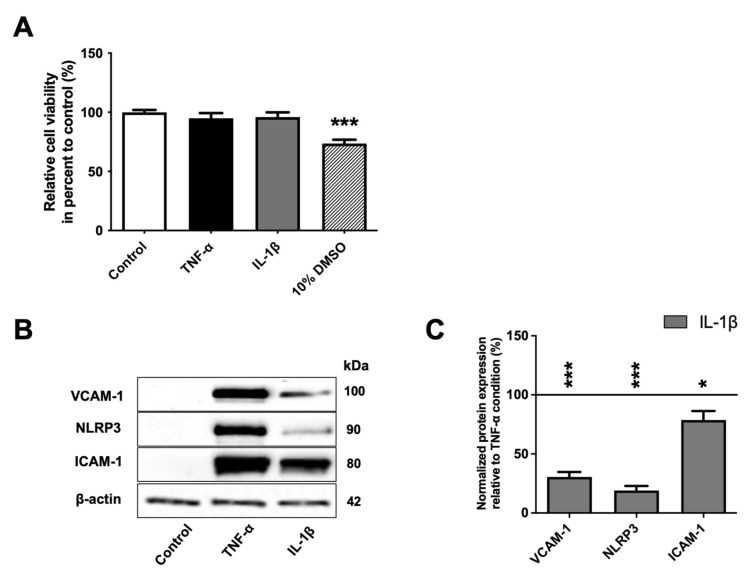
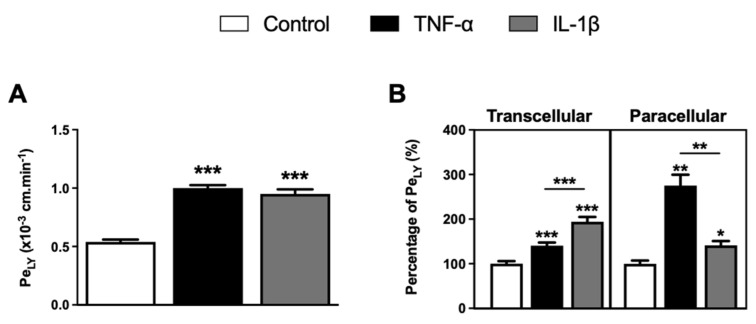
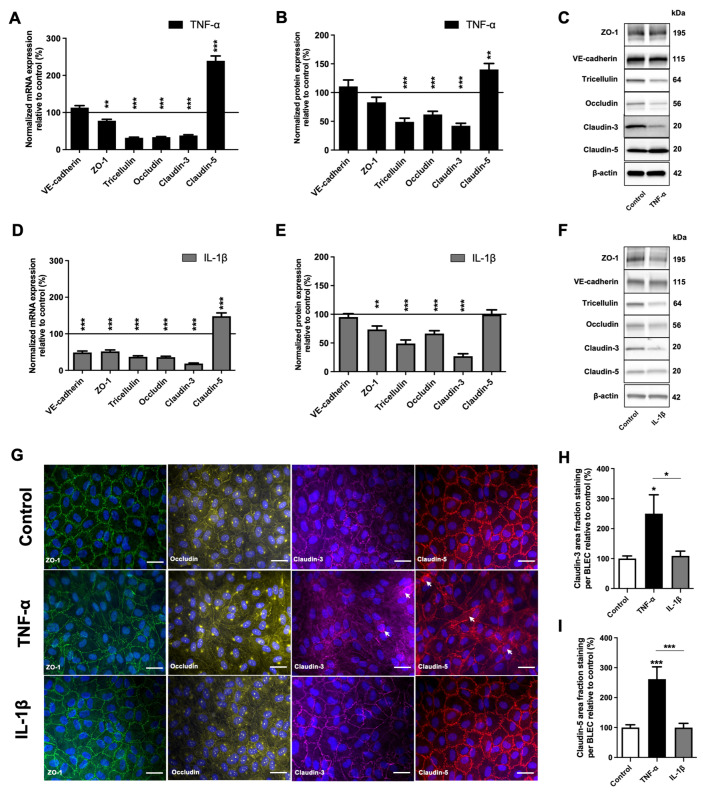
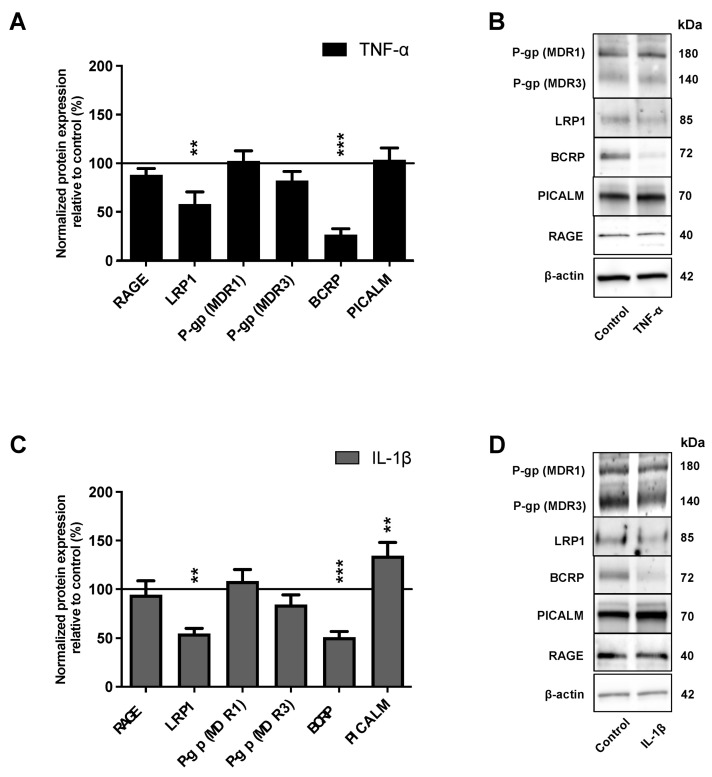
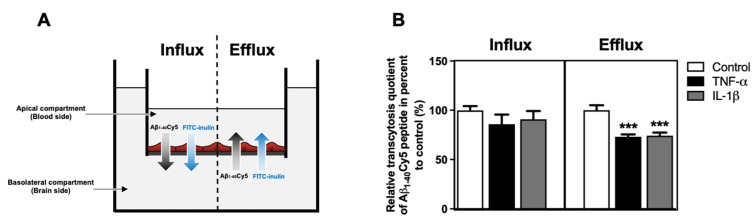
The blood-brain barrier (BBB) is a selective barrier and a functional gatekeeper for the central nervous system (CNS), essential for maintaining brain homeostasis. The BBB is composed of specialized brain endothelial cells (BECs) lining the brain capillaries. The tight junctions formed by BECs regulate paracellular transport, whereas transcellular transport is regulated by specialized transporters, pumps and receptors. Cytokine-induced neuroinflammation, such as the tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), appear to play a role in BBB dysfunction and contribute to the progression of Alzheimer's disease (AD) by contributing to amyloid-β (Aβ) peptide accumulation. Here, we investigated whether TNF-α and IL-1β modulate the permeability of the BBB and alter Aβ peptide transport across BECs. We used a human BBB in vitro model based on the use of brain-like endothelial cells (BLECs) obtained from endothelial cells derived from CD34+ stem cells cocultivated with brain pericytes. We demonstrated that TNF-α and IL-1β differentially induced changes in BLECs' permeability by inducing alterations in the organization of junctional complexes as well as in transcelluar trafficking. Further, TNF-α and IL-1β act directly on BLECs by decreasing LRP1 and BCRP protein expression as well as the specific efflux of Aβ peptide. These results provide mechanisms by which CNS inflammation might modulate BBB permeability and promote Aβ peptide accumulation. A future therapeutic intervention targeting vascular inflammation at the BBB may have the therapeutic potential to slow down the progression of AD.
Keywords: Alzheimer’s disease; IL-1β; TNF-α; amyloid-β peptide; blood-brain barrier; inflammation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Candela P., Gosselet F., Saint-Pol J., Sevin E., Boucau M.C., Boulanger E., Cecchelli R., Fenart L. Apical-to-Basolateral Transport of Amyloid-β Peptides through Blood-Brain Barrier Cells Is Mediated by the Receptor for Advanced Glycation End-Products and Is Restricted by P-Glycoprotein. J. Alzheimer’s Dis. 2010;22:849–859. doi: 10.3233/JAD-2010-100462. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
