

Volatile Dimethyl Disulfide from Guava Plants Regulate Developmental Performance of Asian Citrus Psyllid through Activation of Defense Responses in Neighboring Orange Plants
- PMID: 36142192
- PMCID: PMC9499464
- DOI: 10.3390/ijms231810271
Volatile Dimethyl Disulfide from Guava Plants Regulate Developmental Performance of Asian Citrus Psyllid through Activation of Defense Responses in Neighboring Orange Plants
Abstract
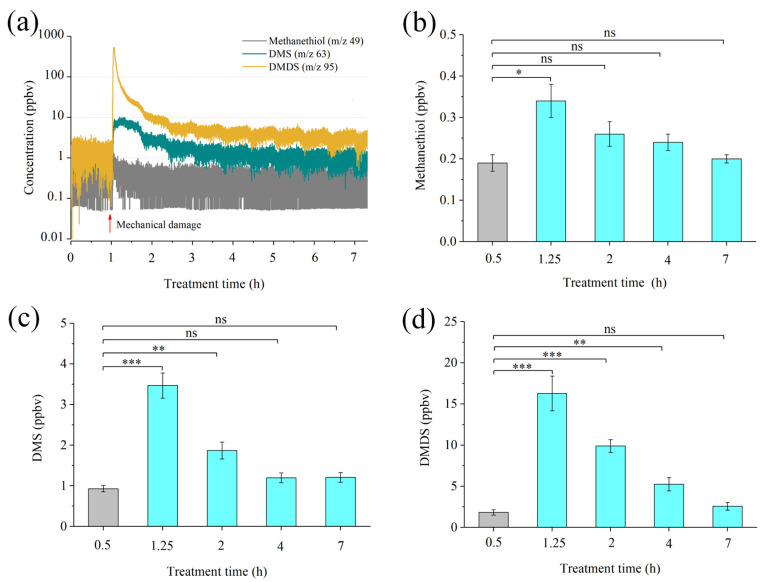
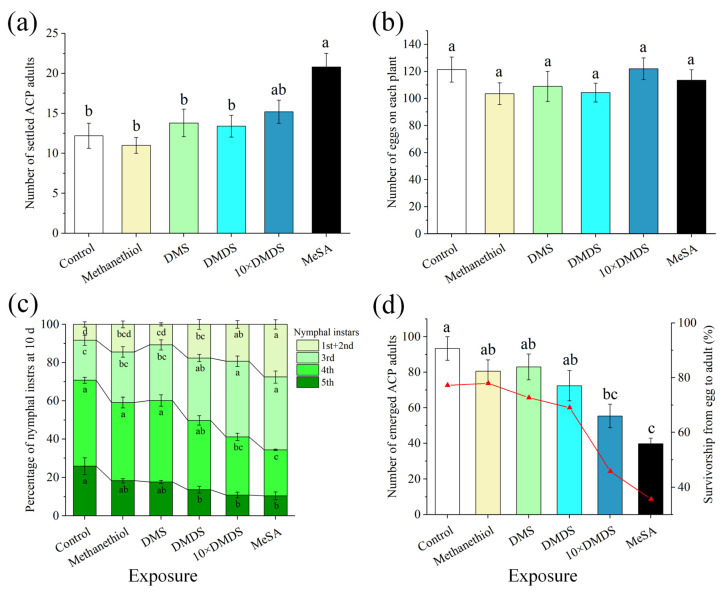
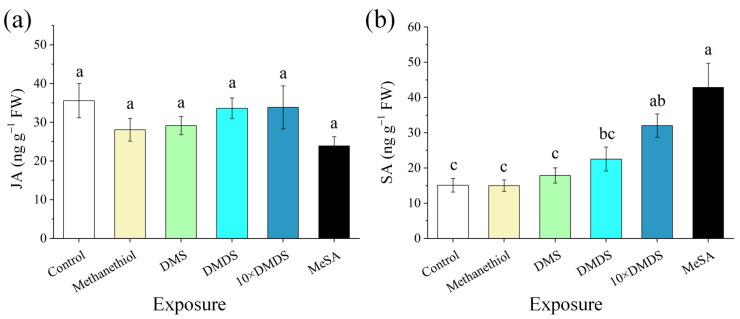
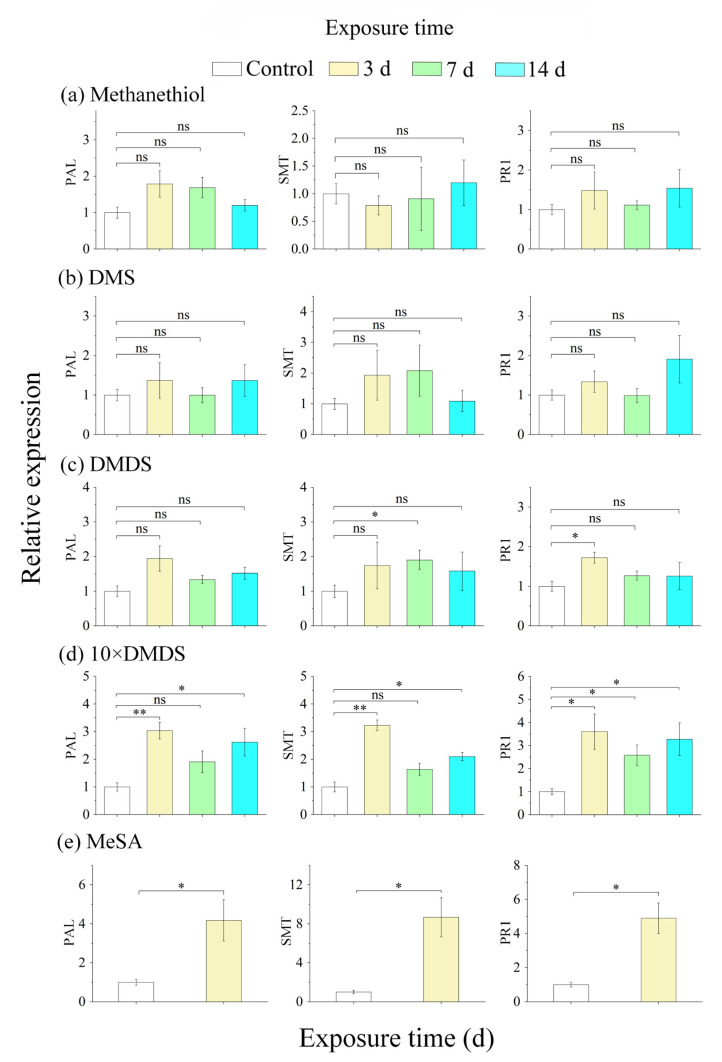
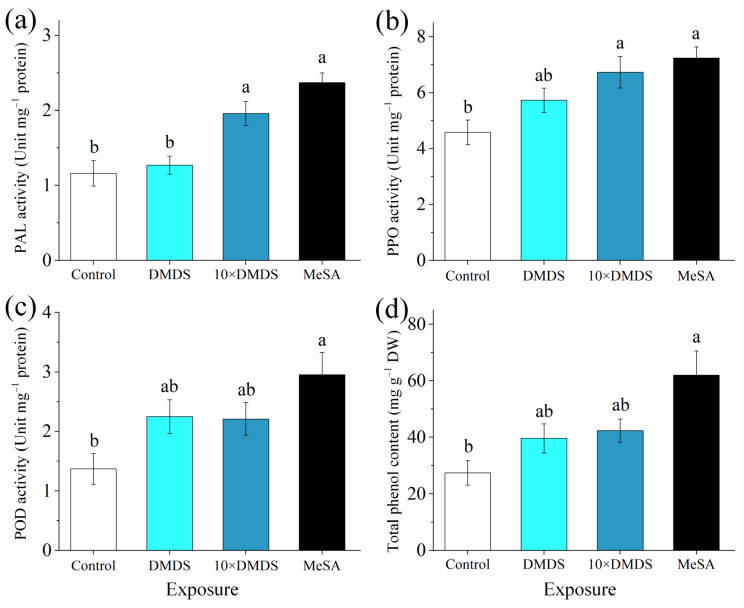
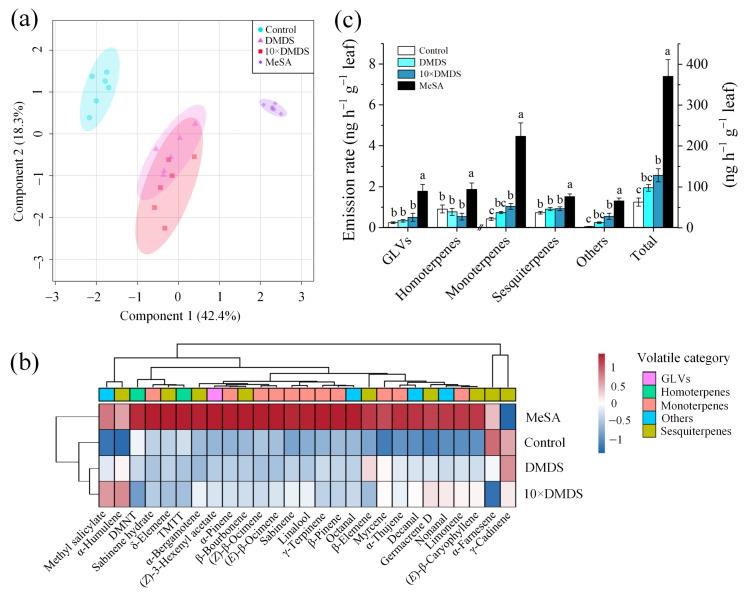
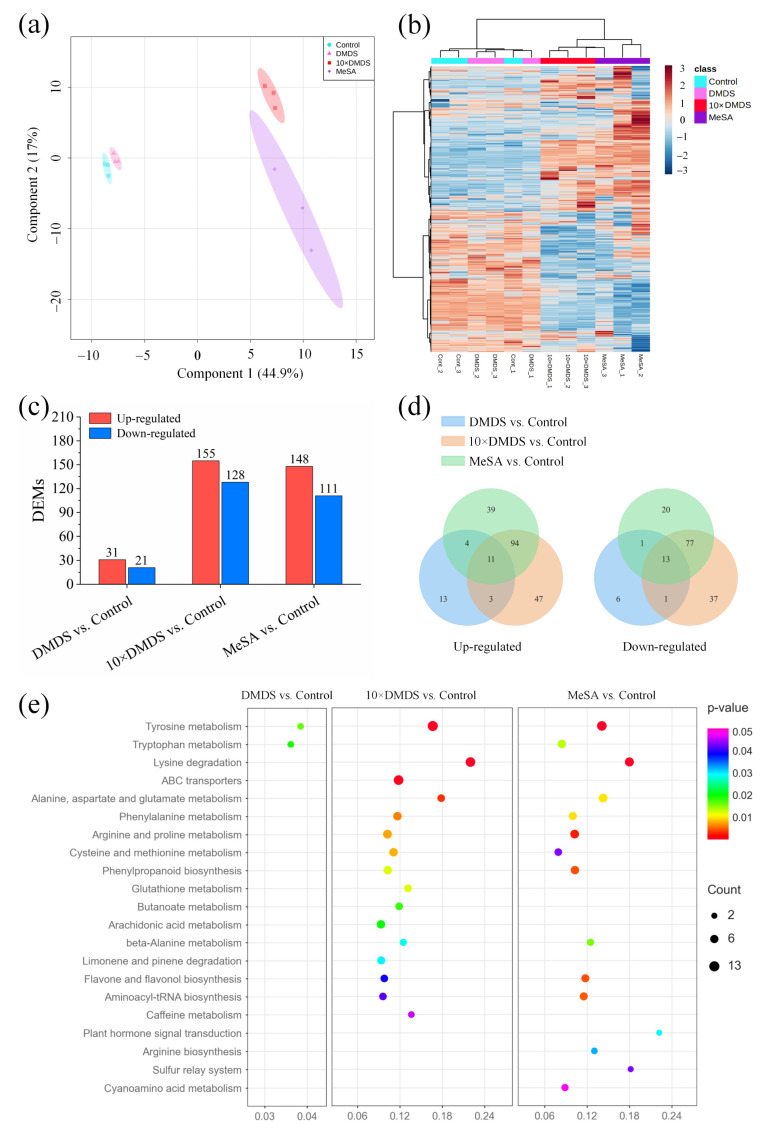
Intercropping with guava (Psidium guajava L.) can assist with the management of Asian citrus psyllid (ACP, Diaphorina citri Kuwayama), the insect vector of the huanglongbing pathogen, in citrus orchards. Sulfur volatiles have a repellent activity and physiological effects, as well as being important components of guava volatiles. In this study, we tested whether the sulfur volatiles emitted by guava plants play a role in plant-plant communications and trigger anti-herbivore activities against ACP in sweet orange plants (Citrus sinensis L. Osbeck). Real-time determination using a proton-transfer-reaction mass spectrometer (PTR-MS) showed that guava plants continuously release methanethiol, dimethyl sulfide (DMS), and dimethyl disulfide (DMDS), and the contents increased rapidly after mechanical damage. The exposure of orange plants to DMDS resulted in the suppression of the developmental performance of ACP. The differential elevation of salicylic acid (SA) levels; the expression of phenylalanine ammonia lyase (PAL), salicylate-O-methyl transferase (SMT), and pathogenesis-related (PR1) genes; the activities of defense-related enzymes PAL, polyphenol oxidase (PPO), and peroxidase (POD); and the total polyphenol content were observed in DMDS-exposed orange plants. The emission of volatiles including myrcene, nonanal, decanal, and methyl salicylate (MeSA) was increased. In addition, phenylpropanoid and flavonoid biosynthesis, and aromatic amino acid (such as phenylalanine, tyrosine, and tryptophan) metabolic pathways were induced. Altogether, our results indicated that DMDS from guava plants can activate defense responses in eavesdropping orange plants and boost their herbivore resistance to ACP, which suggests the possibility of using DMDS as a novel approach for the management of ACP in citrus orchards.
Keywords: Asian citrus psyllid; defense response; dimethyl disulfide; eavesdropping; guava; sulfur volatiles; sweet orange.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Volatile Signals From Guava Plants Prime Defense Signaling and Increase Jasmonate-Dependent Herbivore Resistance in Neighboring Citrus Plants.Front Plant Sci. 2022 Mar 10;13:833562. doi: 10.3389/fpls.2022.833562. eCollection 2022. Front Plant Sci. 2022. PMID: 35371180 Free PMC article.
-
Sulfur volatiles in guava (Psidium guajava L.) leaves: possible defense mechanism.J Agric Food Chem. 2008 Oct 8;56(19):8905-10. doi: 10.1021/jf801735v. Epub 2008 Sep 9. J Agric Food Chem. 2008. PMID: 18778077
-
Exogenous application of the plant signalers methyl jasmonate and salicylic acid induces changes in volatile emissions from citrus foliage and influences the aggregation behavior of Asian citrus psyllid (Diaphorina citri), vector of Huanglongbing.PLoS One. 2018 Mar 29;13(3):e0193724. doi: 10.1371/journal.pone.0193724. eCollection 2018. PLoS One. 2018. PMID: 29596451 Free PMC article.
-
The push-pull strategy for citrus psyllid control.Pest Manag Sci. 2015 Jul;71(7):893-6. doi: 10.1002/ps.3915. Epub 2014 Nov 10. Pest Manag Sci. 2015. PMID: 25256398 Review.
-
Integrated Pest Management Strategies for Asian Citrus Psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae) and Huanglongbing in Citrus for Sarawak, East Malaysia, Borneo.Insects. 2022 Oct 20;13(10):960. doi: 10.3390/insects13100960. Insects. 2022. PMID: 36292909 Free PMC article. Review.
Cited by
-
Volatiles from Pseudomonas palleroniana Strain B-BH16-1 Suppress Aflatoxin Production and Growth of Aspergillus flavus on Coix lacryma-jobi during Storage.Toxins (Basel). 2023 Jan 14;15(1):77. doi: 10.3390/toxins15010077. Toxins (Basel). 2023. PMID: 36668896 Free PMC article.
-
Recent Advances in Plant-Insect Interactions.Int J Mol Sci. 2023 Jul 12;24(14):11338. doi: 10.3390/ijms241411338. Int J Mol Sci. 2023. PMID: 37511097 Free PMC article.
-
Protective enzyme activity regulation in cotton (Gossypium hirsutum L.) in response to Scirpus planiculmis stress.Front Plant Sci. 2022 Nov 28;13:1068419. doi: 10.3389/fpls.2022.1068419. eCollection 2022. Front Plant Sci. 2022. PMID: 36518496 Free PMC article.
-
Volatile dimethyl disulphide emission from Burkholderia cepacia LS-044 suppresses metabolism and budding in caspofungin-resistant Nakaseomyces glabratus NT2.Sci Rep. 2025 Jul 2;15(1):23307. doi: 10.1038/s41598-025-05383-5. Sci Rep. 2025. PMID: 40603923 Free PMC article.
-
Identification and analysis of VOCs released by Rhodococcus ruber GXMZU2400 to promote plant growth and inhibit pathogen growth.BMC Plant Biol. 2025 Apr 29;25(1):559. doi: 10.1186/s12870-025-06582-y. BMC Plant Biol. 2025. PMID: 40301703 Free PMC article.
References
-
- Ninkovic V., Markovic D., Dahlin I. Decoding neighbour volatiles in preparation for future competition and implications for tritrophic interactions. Perspect. Plant Ecol. Evol. Syst. 2016;23:11–17. doi: 10.1016/j.ppees.2016.09.005. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
