

Thebromine Targets Adenosine Receptors to Control Hippocampal Neuronal Function and Damage
- PMID: 36142422
- PMCID: PMC9502181
- DOI: 10.3390/ijms231810510
Thebromine Targets Adenosine Receptors to Control Hippocampal Neuronal Function and Damage
Abstract
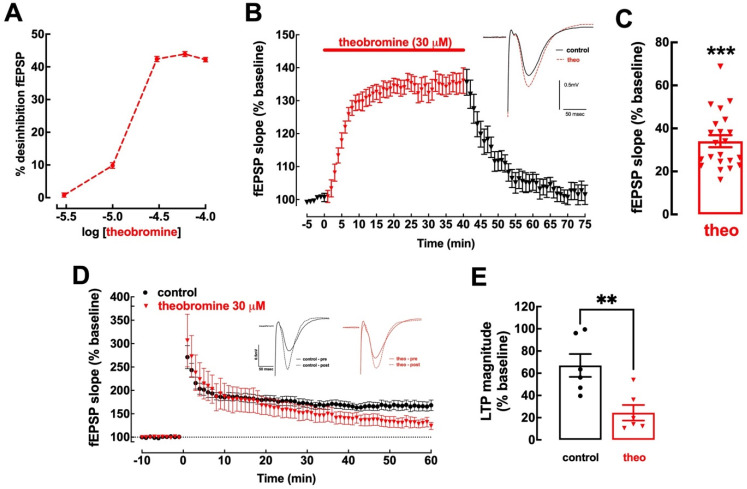
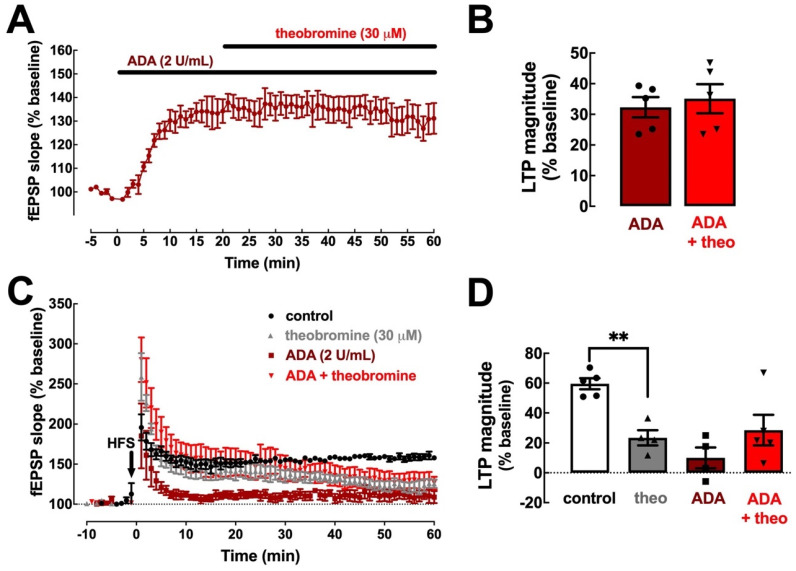
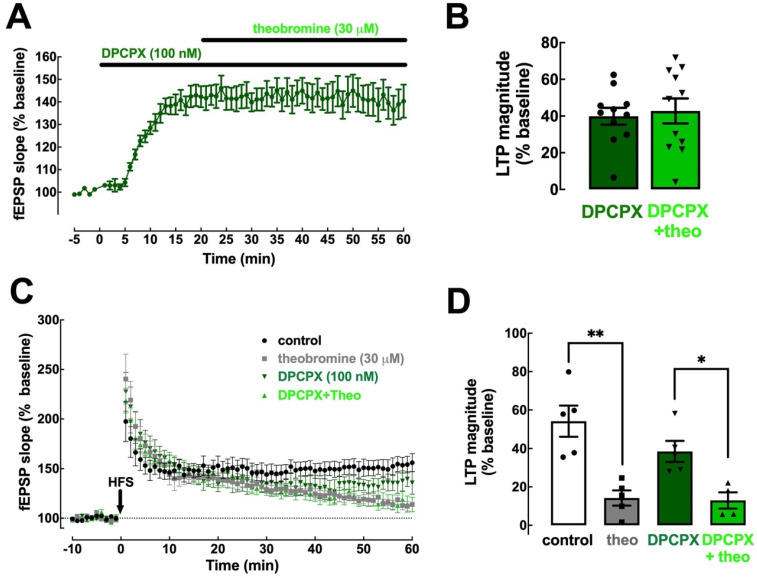
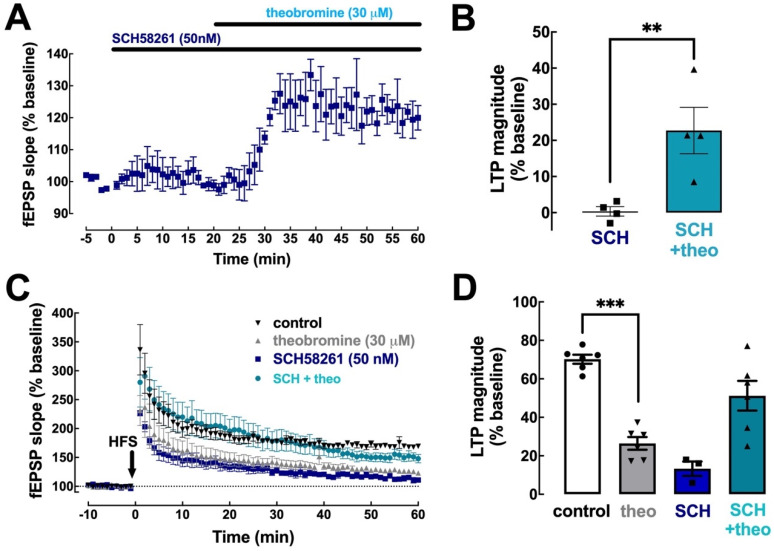
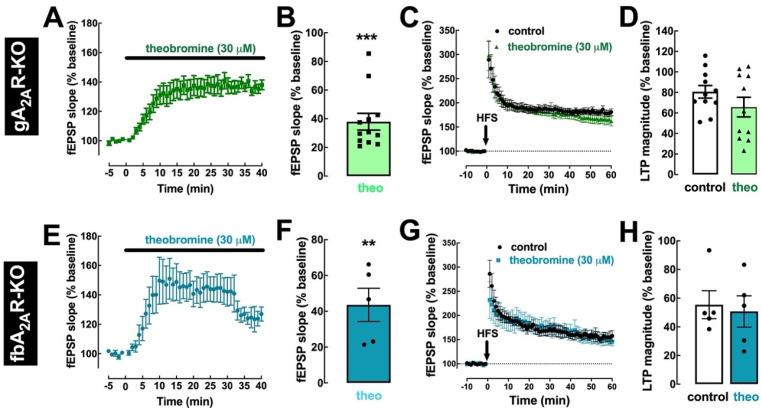
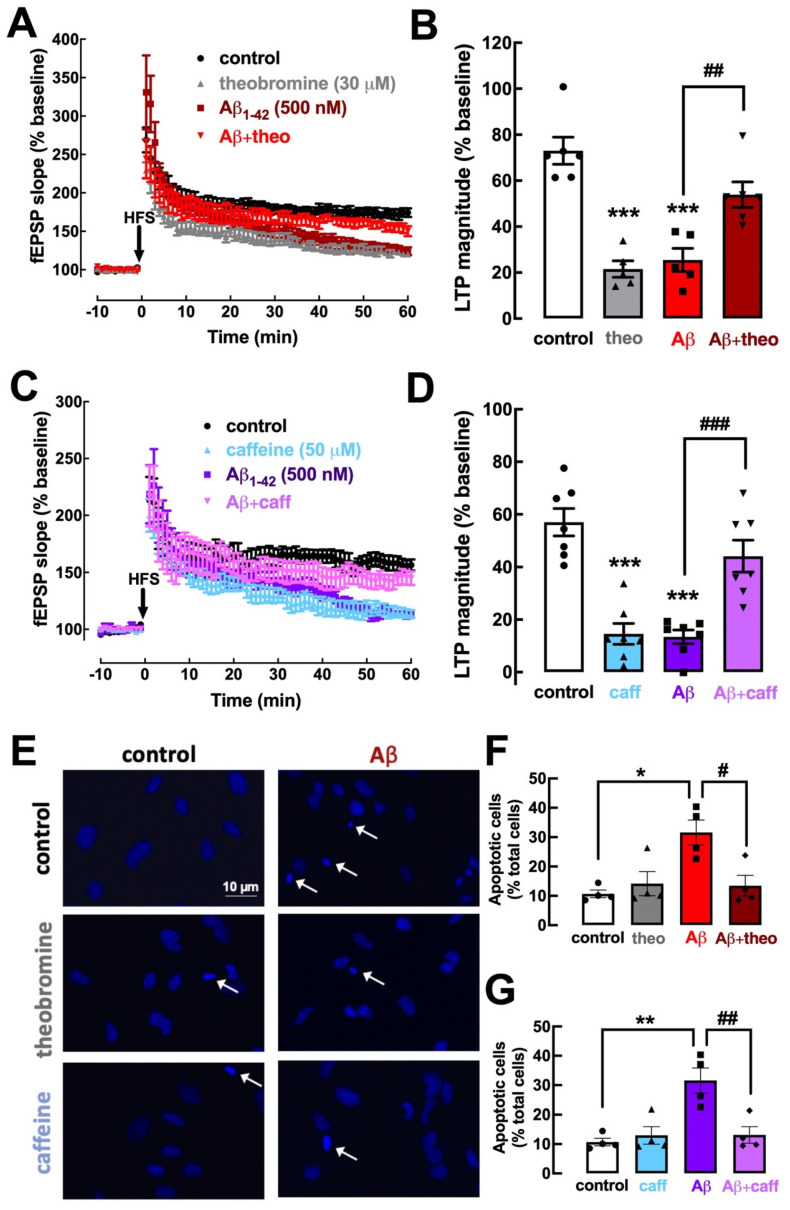
Theobromine is a caffeine metabolite most abundant in dark chocolate, of which consumption is linked with a lower risk of cognitive decline. However, the mechanisms through which theobromine affects neuronal function remain ill-defined. Using electrophysiological recordings in mouse hippocampal synapses, we now characterized the impact of a realistic concentration of theobromine on synaptic transmission and plasticity. Theobromine (30 μM) facilitated synaptic transmission while decreasing the magnitude of long-term potentiation (LTP), with both effects being blunted by adenosine deaminase (2 U/mL). The pharmacological blockade of A1R with DPCPX (100 nM) eliminated the theobromine-dependent facilitation of synaptic transmission, whereas the A2AR antagonist SCH58261 (50 nM), as well as the genetic deletion of A2AR, abrogated the theobromine-induced impairment of LTP. Furthermore, theobromine prevented LTP deficits and neuronal loss, respectively, in mouse hippocampal slices and neuronal cultures exposed to Aβ1-42 peptides, considered a culprit of Alzheimer's disease. Overall, these results indicate that theobromine affects information flow via the antagonism of adenosine receptors, normalizing synaptic plasticity and affording neuroprotection in dementia-related conditions in a manner similar to caffeine.
Keywords: Alzheimer’s disease; adenosine receptors; caffeine; synaptic plasticity; synaptic transmission; theobromine.
Conflict of interest statement
R.A.C. is a scientific advisor of the Institute for Scientific Information on Coffee (ISIC). All other authors declare no competing financial interests.
Figures
Similar articles
-
The physiological effects of caffeine on synaptic transmission and plasticity in the mouse hippocampus selectively depend on adenosine A1 and A2A receptors.Biochem Pharmacol. 2019 Aug;166:313-321. doi: 10.1016/j.bcp.2019.06.008. Epub 2019 Jun 12. Biochem Pharmacol. 2019. PMID: 31199895
-
Adenosine A2A Receptors Shut Down Adenosine A1 Receptor-Mediated Presynaptic Inhibition to Promote Implementation of Hippocampal Long-Term Potentiation.Biomolecules. 2023 Apr 21;13(4):715. doi: 10.3390/biom13040715. Biomolecules. 2023. PMID: 37189461 Free PMC article.
-
Adenosine A2A Receptor Up-Regulation Pre-Dates Deficits of Synaptic Plasticity and of Memory in Mice Exposed to Aβ1-42 to Model Early Alzheimer's Disease.Biomolecules. 2023 Jul 28;13(8):1173. doi: 10.3390/biom13081173. Biomolecules. 2023. PMID: 37627238 Free PMC article.
-
Caffeine, adenosine receptors, and synaptic plasticity.J Alzheimers Dis. 2010;20 Suppl 1:S25-34. doi: 10.3233/JAD-2010-091384. J Alzheimers Dis. 2010. PMID: 20182030 Review.
-
Aquaporin-4 water channels and synaptic plasticity in the hippocampus.Neurochem Int. 2013 Dec;63(7):702-11. doi: 10.1016/j.neuint.2013.05.003. Epub 2013 May 15. Neurochem Int. 2013. PMID: 23684954 Free PMC article. Review.
Cited by
-
Urinary Metabolic Profiling During Epileptogenesis in Rat Model of Lithium-Pilocarpine-Induced Temporal Lobe Epilepsy.Biomedicines. 2025 Feb 27;13(3):588. doi: 10.3390/biomedicines13030588. Biomedicines. 2025. PMID: 40149565 Free PMC article.
-
Caffeine is negatively associated with depression in patients aged 20 and older.Front Psychiatry. 2022 Dec 1;13:1037579. doi: 10.3389/fpsyt.2022.1037579. eCollection 2022. Front Psychiatry. 2022. PMID: 36532163 Free PMC article.
-
Targeting protein aggregation using a cocoa-bean shell extract to reduce α-synuclein toxicity in models of Parkinson's disease.Curr Res Food Sci. 2024 Oct 24;9:100888. doi: 10.1016/j.crfs.2024.100888. eCollection 2024. Curr Res Food Sci. 2024. PMID: 39525389 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
