

Extended N-Terminal Acetyltransferase Naa50 in Filamentous Fungi Adds to Naa50 Diversity
- PMID: 36142717
- PMCID: PMC9500918
- DOI: 10.3390/ijms231810805
Extended N-Terminal Acetyltransferase Naa50 in Filamentous Fungi Adds to Naa50 Diversity
Abstract
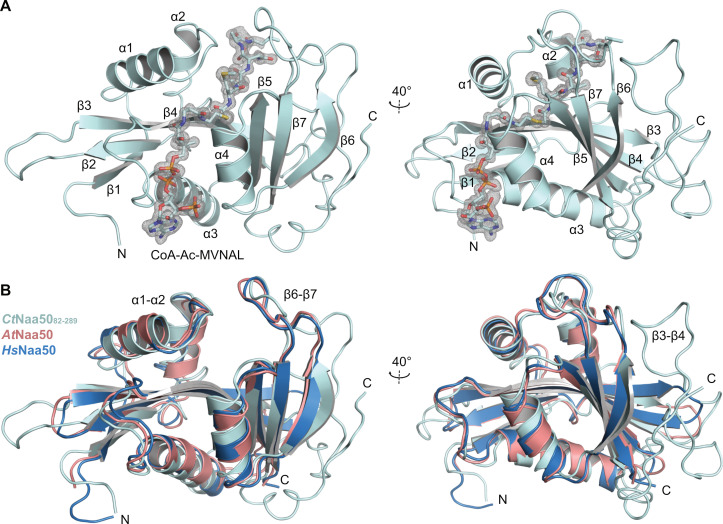
Most eukaryotic proteins are N-terminally acetylated by a set of Nα acetyltransferases (NATs). This ancient and ubiquitous modification plays a fundamental role in protein homeostasis, while mutations are linked to human diseases and phenotypic defects. In particular, Naa50 features species-specific differences, as it is inactive in yeast but active in higher eukaryotes. Together with NatA, it engages in NatE complex formation for cotranslational acetylation. Here, we report Naa50 homologs from the filamentous fungi Chaetomium thermophilum and Neurospora crassa with significant N- and C-terminal extensions to the conserved GNAT domain. Structural and biochemical analyses show that CtNaa50 shares the GNAT structure and substrate specificity with other homologs. However, in contrast to previously analyzed Naa50 proteins, it does not form NatE. The elongated N-terminus increases Naa50 thermostability and binds to dynein light chain protein 1, while our data suggest that conserved positive patches in the C-terminus allow for ribosome binding independent of NatA. Our study provides new insights into the many facets of Naa50 and highlights the diversification of NATs during evolution.
Keywords: Chaetomium thermophilum; GNAT domain; N-terminal acetyltransferase; NAT; Naa50; NatE; Neurospora crassa; X-ray structure; dynein light chain protein 1; ribosome association.
Conflict of interest statement
The authors declare no conflict of interest.
Figures










References
-
- Arnesen T., Van Damme P., Polevoda B., Helsens K., Evjenth R., Colaert N., Varhaug J.E., Vandekerckhove J., Lillehaug J.R., Sherman F., et al. Proteomics Analyses Reveal the Evolutionary Conservation and Divergence of N-Terminal Acetyltransferases from Yeast and Humans. Proc. Natl. Acad. Sci. USA. 2009;106:8157–8162. doi: 10.1073/pnas.0901931106. - DOI - PMC - PubMed
-
- Bienvenut W.V., Sumpton D., Martinez A., Lilla S., Espagne C., Meinnel T., Giglione C. Comparative Large Scale Characterization of Plant versus Mammal Proteins Reveals Similar and Idiosyncratic N-alpha-Acetylation Features. Mol. Cell. Proteom. 2012;11:M111.015131. doi: 10.1074/mcp.M111.015131. - DOI - PMC - PubMed
-
- Lange P.F., Huesgen P.F., Nguyen K., Overall C.M. Annotating N termini for the human proteome project: N termini and Nalpha-acetylation status differentiate stable cleaved protein species from degradation remnants in the human erythrocyte proteome. J. Proteome Res. 2014;13:2028–2044. doi: 10.1021/pr401191w. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
