

Bile Salt Hydrolases with Extended Substrate Specificity Confer a High Level of Resistance to Bile Toxicity on Atopobiaceae Bacteria
- PMID: 36142891
- PMCID: PMC9506489
- DOI: 10.3390/ijms231810980
Bile Salt Hydrolases with Extended Substrate Specificity Confer a High Level of Resistance to Bile Toxicity on Atopobiaceae Bacteria
Abstract
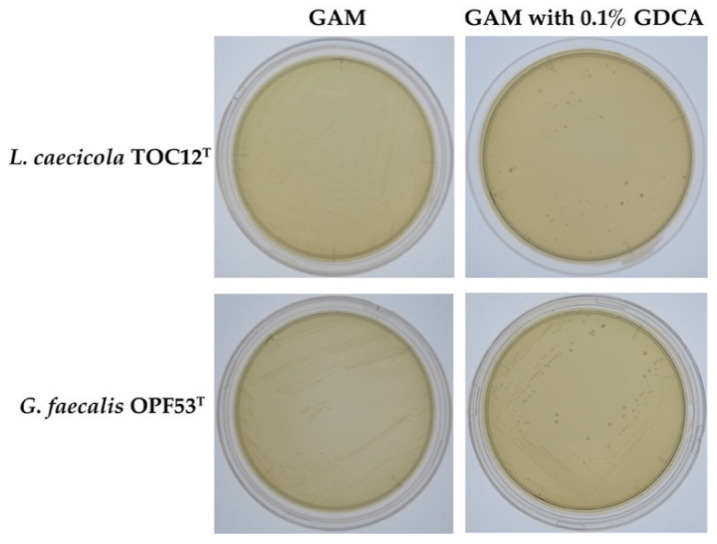
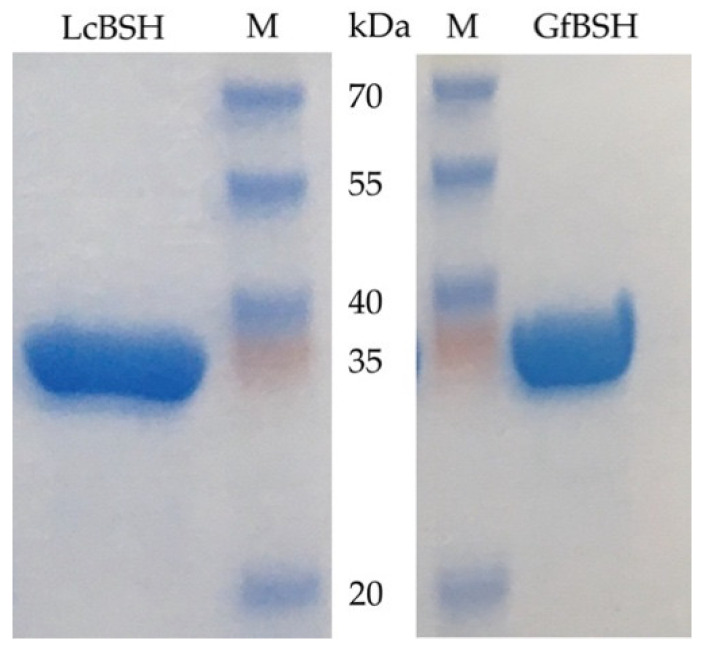
The bile resistance of intestinal bacteria is among the key factors responsible for their successful colonization of and survival in the mammalian gastrointestinal tract. In this study, we demonstrated that lactate-producing Atopobiaceae bacteria (Leptogranulimonas caecicola TOC12T and Granulimonas faecalis OPF53T) isolated from mouse intestine showed high resistance to mammalian bile extracts, due to significant bile salt hydrolase (BSH) activity. We further succeeded in isolating BSH proteins (designated LcBSH and GfBSH) from L. caecicola TOC12T and G. faecalis OPF53T, respectively, and characterized their enzymatic features. Interestingly, recombinant LcBSH and GfBSH proteins exhibited BSH activity against 12 conjugated bile salts, indicating that LcBSH and GfBSH have much broader substrate specificity than the previously identified BSHs from lactic acid bacteria, which are generally known to hydrolyze six bile salt isomers. Phylogenetic analysis showed that LcBSH and GfBSH had no affinities with any known BSH subgroup and constituted a new BSH subgroup in the phylogeny. In summary, we discovered functional BSHs with broad substrate specificity from Atopobiaceae bacteria and demonstrated that these BSH enzymes confer bile resistance to L. caecicola TOC12T and G. faecalis OPF53T.
Keywords: Granulimonas faecalis; Leptogranulimonas caecicola; bile resistance; bile salt hydrolase; probiotics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Granulimonas faecalis gen. nov., sp. nov., and Leptogranulimonas caecicola gen. nov., sp. nov., novel lactate-producing Atopobiaceae bacteria isolated from mouse intestines, and an emended description of the family Atopobiaceae.Int J Syst Evol Microbiol. 2022 Oct;72(10). doi: 10.1099/ijsem.0.005596. Int J Syst Evol Microbiol. 2022. PMID: 36288087
-
Evidence for the Worldwide Distribution of a Bile Salt Hydrolase Gene in Enterococcus faecium Through Horizontal Gene Transfer.Int J Mol Sci. 2025 Jan 13;26(2):612. doi: 10.3390/ijms26020612. Int J Mol Sci. 2025. PMID: 39859326 Free PMC article.
-
Bile salt hydrolase activity is present in nonintestinal lactic acid bacteria at an intermediate level.Appl Microbiol Biotechnol. 2019 Jan;103(2):893-902. doi: 10.1007/s00253-018-9492-5. Epub 2018 Nov 12. Appl Microbiol Biotechnol. 2019. PMID: 30421106
-
Molecular features of bile salt hydrolases and relevance in human health.Biochim Biophys Acta Gen Subj. 2017 Jan;1861(1 Pt A):2981-2991. doi: 10.1016/j.bbagen.2016.09.024. Epub 2016 Sep 25. Biochim Biophys Acta Gen Subj. 2017. PMID: 27681686 Review.
-
New insights into bacterial bile resistance mechanisms: the role of bile salt hydrolase and its impact on human health.Food Res Int. 2018 Oct;112:250-262. doi: 10.1016/j.foodres.2018.06.035. Epub 2018 Jun 20. Food Res Int. 2018. PMID: 30131136 Review.
Cited by
-
Spaceflight alters host-gut microbiota interactions.NPJ Biofilms Microbiomes. 2024 Aug 29;10(1):71. doi: 10.1038/s41522-024-00545-1. NPJ Biofilms Microbiomes. 2024. PMID: 39209868 Free PMC article.
-
Christensenella minuta interacts with multiple gut bacteria.Front Microbiol. 2024 Feb 19;15:1301073. doi: 10.3389/fmicb.2024.1301073. eCollection 2024. Front Microbiol. 2024. PMID: 38440147 Free PMC article.
-
Oral exposure to benzalkonium chlorides in male and female mice reveals alteration of the gut microbiome and bile acid profile.Toxicol Sci. 2024 Dec 1;202(2):265-277. doi: 10.1093/toxsci/kfae116. Toxicol Sci. 2024. PMID: 39363503
-
Prebiotic potential of green banana flour: impact on gut microbiota modulation and microbial metabolic activity in a murine model.Front Nutr. 2023 Oct 31;10:1249358. doi: 10.3389/fnut.2023.1249358. eCollection 2023. Front Nutr. 2023. PMID: 38024360 Free PMC article.
-
Gut Microbiota Dysbiosis in Diabetic Retinopathy-Current Knowledge and Future Therapeutic Targets.Life (Basel). 2023 Apr 7;13(4):968. doi: 10.3390/life13040968. Life (Basel). 2023. PMID: 37109497 Free PMC article. Review.
References
-
- Fuller R. Probiotics. Springer; Berlin, Germany: 1992. History and development of probiotics; pp. 1–8.
-
- Kadooka Y., Sato M., Imaizumi K., Ogawa A., Ikuyama K., Akai Y., Okano M., Kagoshima M., Tsuchida T. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur. J. Clin. Nutr. 2010;64:636–643. doi: 10.1038/ejcn.2010.19. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
