

How to Shut Down Transcription in Archaea during Virus Infection
- PMID: 36144426
- PMCID: PMC9501531
- DOI: 10.3390/microorganisms10091824
How to Shut Down Transcription in Archaea during Virus Infection
Abstract
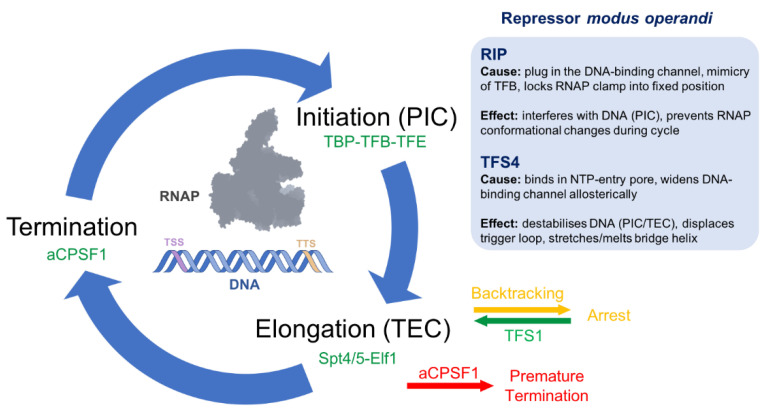
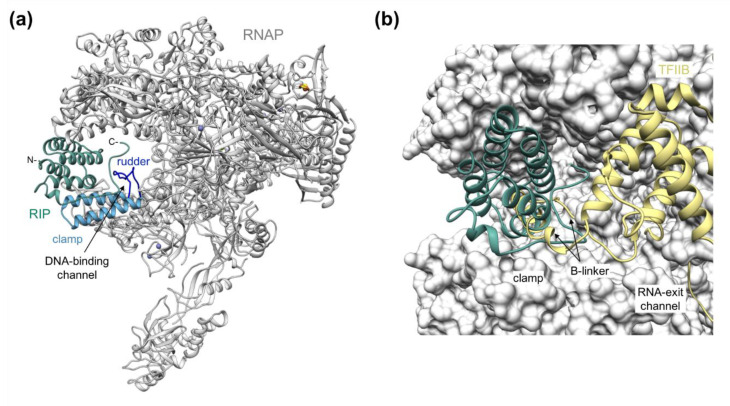
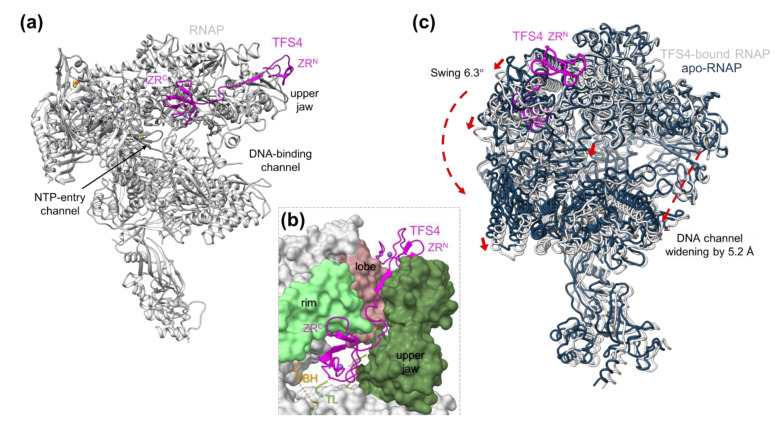
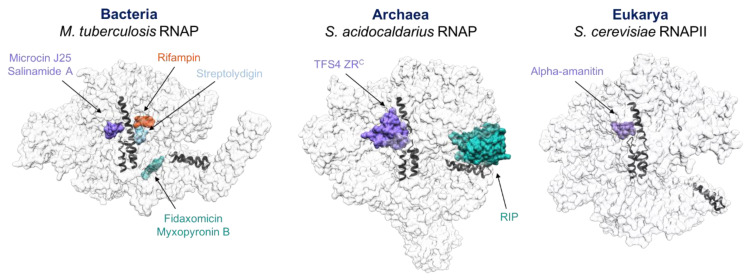
Multisubunit RNA polymerases (RNAPs) carry out transcription in all domains of life; during virus infection, RNAPs are targeted by transcription factors encoded by either the cell or the virus, resulting in the global repression of transcription with distinct outcomes for different host-virus combinations. These repressors serve as versatile molecular probes to study RNAP mechanisms, as well as aid the exploration of druggable sites for the development of new antibiotics. Here, we review the mechanisms and structural basis of RNAP inhibition by the viral repressor RIP and the crenarchaeal negative regulator TFS4, which follow distinct strategies. RIP operates by occluding the DNA-binding channel and mimicking the initiation factor TFB/TFIIB. RIP binds tightly to the clamp and locks it into one fixed position, thereby preventing conformational oscillations that are critical for RNAP function as it progresses through the transcription cycle. TFS4 engages with RNAP in a similar manner to transcript cleavage factors such as TFS/TFIIS through the NTP-entry channel; TFS4 interferes with the trigger loop and bridge helix within the active site by occlusion and allosteric mechanisms, respectively. The conformational changes in RNAP described above are universally conserved and are also seen in inactive dimers of eukaryotic RNAPI and several inhibited RNAP complexes of both bacterial and eukaryotic RNA polymerases, including inactive states that precede transcription termination. A comparison of target sites and inhibitory mechanisms reveals that proteinaceous repressors and RNAP-specific antibiotics use surprisingly common ways to inhibit RNAP function.
Keywords: RNA polymerase; antibiotics; archaea; evolution; transcription inhibition; viruses.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study, in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
Structural basis of RNA polymerase inhibition by viral and host factors.Nat Commun. 2021 Sep 17;12(1):5523. doi: 10.1038/s41467-021-25666-5. Nat Commun. 2021. PMID: 34535646 Free PMC article.
-
The transcript cleavage factor paralogue TFS4 is a potent RNA polymerase inhibitor.Nat Commun. 2017 Dec 4;8(1):1914. doi: 10.1038/s41467-017-02081-3. Nat Commun. 2017. PMID: 29203770 Free PMC article.
-
Prokaryotic sigma factors and their transcriptional counterparts in Archaea and Eukarya.Appl Microbiol Biotechnol. 2020 May;104(10):4289-4302. doi: 10.1007/s00253-020-10577-0. Epub 2020 Mar 30. Appl Microbiol Biotechnol. 2020. PMID: 32232532 Review.
-
Structure and Function of RNA Polymerases and the Transcription Machineries.Subcell Biochem. 2017;83:225-270. doi: 10.1007/978-3-319-46503-6_9. Subcell Biochem. 2017. PMID: 28271479 Review.
-
Hold on!: RNA polymerase interactions with the nascent RNA modulate transcription elongation and termination.RNA Biol. 2010 May-Jun;7(3):310-5. doi: 10.4161/rna.7.3.11912. Epub 2010 May 26. RNA Biol. 2010. PMID: 20473037 Free PMC article. Review.
Cited by
-
Learning from the Invaders: What Viruses Teach Us about RNA-Based Regulation in Microbes.Microorganisms. 2022 Oct 25;10(11):2106. doi: 10.3390/microorganisms10112106. Microorganisms. 2022. PMID: 36363698 Free PMC article.
-
Novel Insights into the Mechanisms of Microbial Transcription and Translation.Microorganisms. 2023 Jun 30;11(7):1720. doi: 10.3390/microorganisms11071720. Microorganisms. 2023. PMID: 37512892 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
