

NAT10: An RNA cytidine transferase regulates fatty acid metabolism in cancer cells
- PMID: 36149760
- PMCID: PMC9505754
- DOI: 10.1002/ctm2.1045
NAT10: An RNA cytidine transferase regulates fatty acid metabolism in cancer cells
Abstract
Background: N-4 cytidine acetylation (ac4C) is an epitranscriptomics modification catalyzed by N-acetyltransferase 10 (NAT10); important for cellular mRNA stability, rRNA biogenesis, cell proliferation and epithelial to mesenchymal transition (EMT). However, whether other crucial pathways are regulated by NAT10-dependent ac4C modification in cancer cells remains unclear. Therefore, in this study, we explored the impact of NAT10 depletion in cancer cells using unbiased RNA-seq.
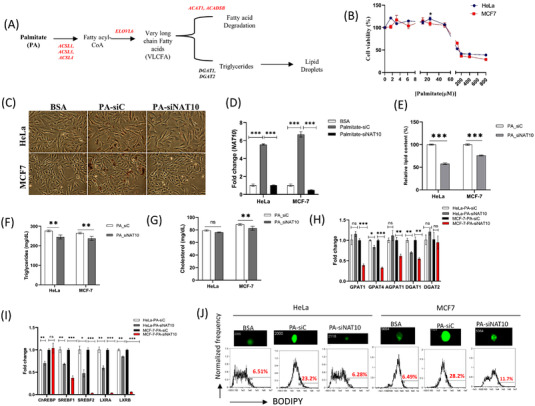
Methods: High-throughput sequencing of knockdown NAT10 in cancer cells was conducted to identify enriched pathways. Acetylated RNA immunoprecipitation-seq (acRIP-seq) and RIP-PCR were used to map and determine ac4C levels of RNA. Exogenous palmitate uptake assay was conducted to assess NAT10 knockdown cancer cells using Oil Red O staining and lipid content analysis. Gas-chromatography-tandem mass spectroscopy (GC/MS) was used to perform untargeted lipidomics.
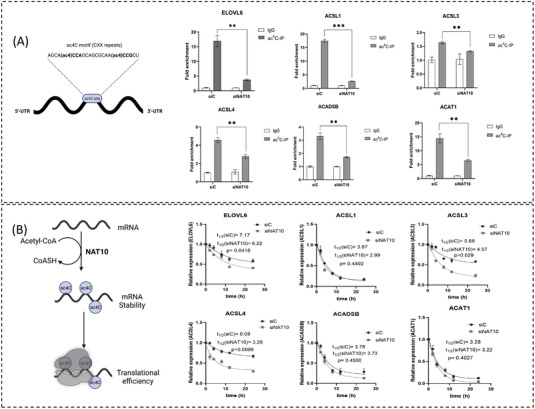
Results: High-throughput sequencing of NAT10 knockdown in cancer cells revealed fatty acid (FA) metabolism as the top enriched pathway through the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis in differentially downregulated genes. FA metabolic genes such as ELOLV6, ACSL1, ACSL3, ACSL4, ACADSB and ACAT1 were shown to be stabilised via NAT10-dependent ac4C RNA acetylation. Additionally, NAT10 depletion was shown to significantly reduce the levels of overall lipid content, triglycerides and total cholesterol. Further, NAT10 depletion in palmitate-loaded cancer cells showed decrease in ac4C levels across the RNA transcripts of FA metabolic genes. In untargeted lipidomics, 496 out of 2 279 lipids were statistically significant in NAT10 depleted cancer cells, of which pathways associated with FA metabolism are the most enriched.
Conclusions: Conclusively, our results provide novel insights into the impact of NAT10-mediated ac4C modification as a crucial regulatory factor during FA metabolism and showed the benefit of targeting NAT10 for cancer treatment.
Keywords: NAT10; ac4C; cancer; fatty acid metabolism.
© 2022 The Authors. Clinical and Translational Medicine published by John Wiley & Sons Australia, Ltd on behalf of Shanghai Institute of Clinical Bioinformatics.
Conflict of interest statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Figures







References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous