

Influence of Zika virus 3'-end sequence and nonstructural protein evolution on the viral replication competence and virulence
- PMID: 36149812
- PMCID: PMC9621255
- DOI: 10.1080/22221751.2022.2128433
Influence of Zika virus 3'-end sequence and nonstructural protein evolution on the viral replication competence and virulence
Abstract
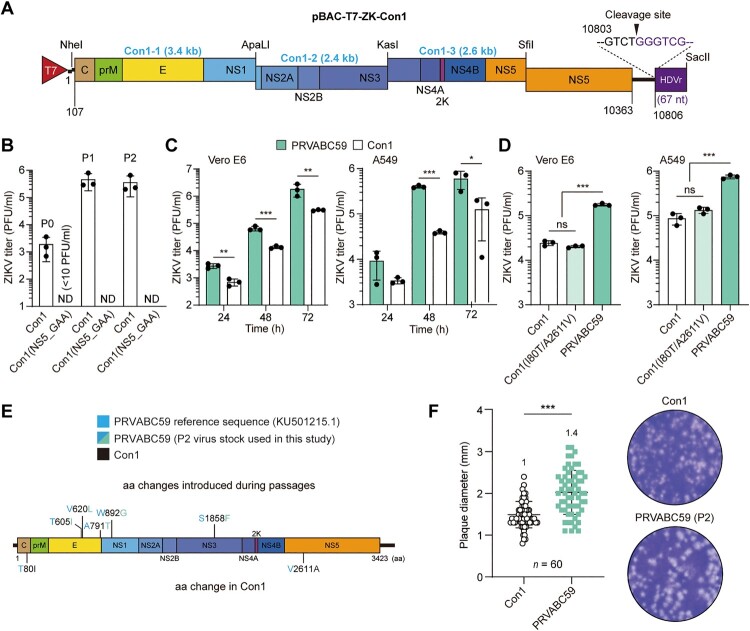
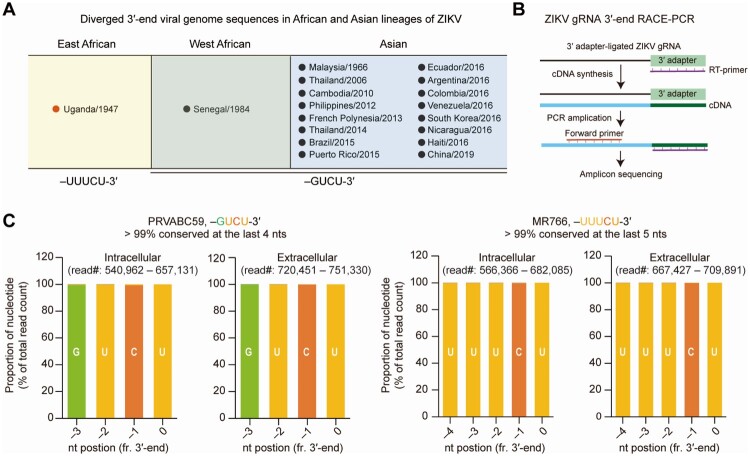
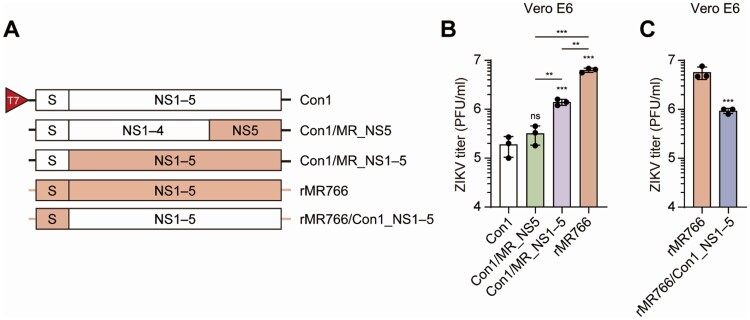
Zika virus (ZIKV) has been circulating in human networks over 70 years since its first appearance in Africa, yet little is known about whether the viral 3'-terminal sequence and nonstructural (NS) protein diverged genetically from ancient ZIKV have different effects on viral replication and virulence in currently prevailing Asian lineage ZIKV. Here we show, by a reverse genetics approach using an infectious cDNA clone for a consensus sequence (Con1) of ZIKV, which represents Asian ZIKV strains, and another clone derived from the MR766 strain isolated in Uganda, Africa in 1947, that the 3'-end sequence -UUUCU-3' homogeneously present in MR766 genome and the -GUCU-3' sequence strictly conserved in Asian ZIKV isolates are functionally equivalent in viral replication and gene expression. By gene swapping experiments using the two infectious cDNA clones, we show that the NS1-5 proteins of MR766 enhance replication competence of ZIKV Con1. The Con1, which was less virulent than MR766, acquired severe bilateral hindlimb paralysis when its NS1-5 genes were replaced by the counterparts of MR766 in type I interferon receptor (IFNAR1)-deficient A129 mice. Moreover, MR766 NS5 RNA-dependent RNA polymerase (RdRp) alone also rendered the Con1 virulent, despite there being no difference in RdRp activity between MR766 and Con1 NS5 proteins. By contrast, the Con1 derivatives expressing MR766 Nsps, like Con1, did not develop severe disease in wild-type mice treated with an IFNAR1 blocking antibody. Together, our findings uncover an unprecedented role for ZIKV NS proteins in determining viral pathogenicity in immunocompromised hosts.
Keywords: 3′-end sequence variants; Zika virus; infectious cDNA clone; nonstructural proteins; virulence determinant.
Conflict of interest statement
J.-W.O., H.-G.J, and H.C. are inventors on a pending patent related to this work filed by University Industry Foundation (UIF), Yonsei University. The remaining authors have no conflicts of interest to declare.
Figures
Similar articles
-
An Alanine-to-Valine Substitution in the Residue 175 of Zika Virus NS2A Protein Affects Viral RNA Synthesis and Attenuates the Virus In Vivo.Viruses. 2018 Oct 7;10(10):547. doi: 10.3390/v10100547. Viruses. 2018. PMID: 30301244 Free PMC article.
-
Rescue of the 1947 Zika Virus Prototype Strain with a Cytomegalovirus Promoter-Driven cDNA Clone.mSphere. 2016 Sep 28;1(5):e00246-16. doi: 10.1128/mSphere.00246-16. eCollection 2016 Sep-Oct. mSphere. 2016. PMID: 27704051 Free PMC article.
-
The NS1 protein of contemporary West African Zika virus potentiates viral replication and reduces innate immune activation.PLoS Negl Trop Dis. 2024 Aug 23;18(8):e0012146. doi: 10.1371/journal.pntd.0012146. eCollection 2024 Aug. PLoS Negl Trop Dis. 2024. PMID: 39178324 Free PMC article.
-
Zika virus: Critical crosstalk between pathogenesis, cytopathic effects, and macroautophagy.J Cell Biochem. 2024 Nov;125(11):e30438. doi: 10.1002/jcb.30438. Epub 2023 Jun 19. J Cell Biochem. 2024. PMID: 37334850 Review.
-
Structure and function of Zika virus NS5 protein: perspectives for drug design.Cell Mol Life Sci. 2018 May;75(10):1723-1736. doi: 10.1007/s00018-018-2751-x. Epub 2018 Feb 8. Cell Mol Life Sci. 2018. PMID: 29423529 Free PMC article. Review.
Cited by
-
Genetically modified ZIKA virus as a microRNA-sensitive oncolytic virus against central nervous system tumors.Mol Ther. 2024 Feb 7;32(2):440-456. doi: 10.1016/j.ymthe.2024.01.006. Epub 2024 Jan 11. Mol Ther. 2024. PMID: 38213031 Free PMC article.
-
Zika virus-induces metabolic alterations in fetal neuronal progenitors that could influence in neurodevelopment during early pregnancy.Biol Open. 2023 Apr 15;12(4):bio059889. doi: 10.1242/bio.059889. Epub 2023 Apr 21. Biol Open. 2023. PMID: 37093064 Free PMC article.
-
Replication properties of a contemporary Zika virus from West Africa.PLoS Negl Trop Dis. 2024 Jul 5;18(7):e0012066. doi: 10.1371/journal.pntd.0012066. eCollection 2024 Jul. PLoS Negl Trop Dis. 2024. PMID: 38968296 Free PMC article.
References
-
- Oehler E, Watrin L, Larre P, et al. . Zika virus infection complicated by Guillain-Barre syndrome - case report, French Polynesia, December 2013. Euro Surveill. 2014;19(9):4–6. - PubMed
-
- Fauci AS, Morens DM.. Zika virus in the Americas - yet another Arbovirus threat. N Engl J Med. 2016;374(7):601–604. - PubMed
-
- Dick GWA, Haddow AJ, Uganda S.. Virus. A hitherto unrecorded virus isolated from mosquitoes in Ugnada. (I). Isolation and pathogenicity. Trans R Soc Trop Med Hyg. 1952;46(6):600–618. - PubMed
-
- Boorman JP, Draper CC.. Isolations of arboviruses in the Lagos area of Nigeria, and a survey of antibodies to them in man and animals. Trans R Soc Trop Med Hyg. 1968;62(2):269–277. - PubMed
-
- Wikan N, Smith DR.. First published report of Zika virus infection in people: Simpson, not MacNamara. Lancet Infect Dis. 2017;17(1):15–17. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous