

Limb, joint and pelvic kinematic control in the quail coping with steps upwards and downwards
- PMID: 36151454
- PMCID: PMC9508109
- DOI: 10.1038/s41598-022-20247-y
Limb, joint and pelvic kinematic control in the quail coping with steps upwards and downwards
Abstract
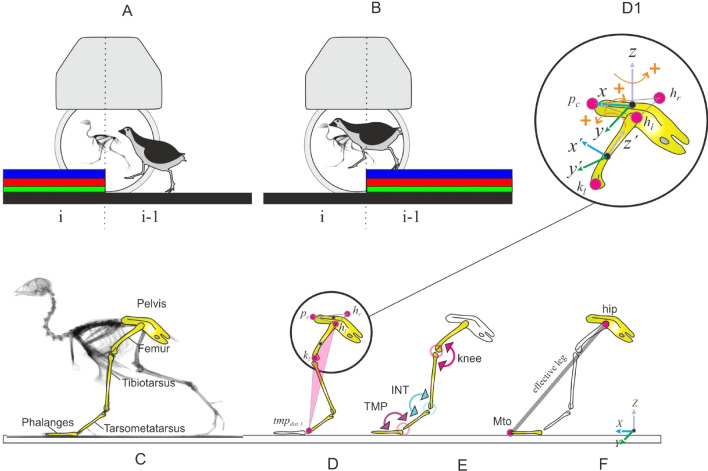
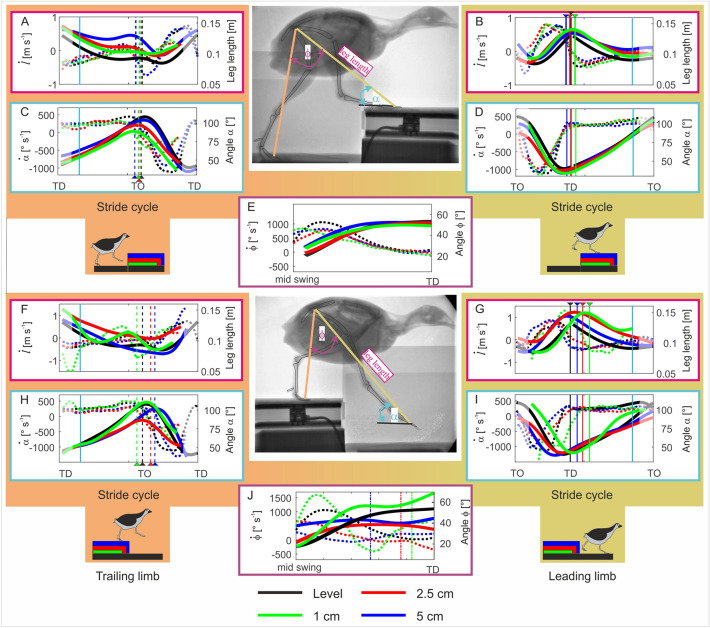
Small cursorial birds display remarkable walking skills and can negotiate complex and unstructured terrains with ease. The neuromechanical control strategies necessary to adapt to these challenging terrains are still not well understood. Here, we analyzed the 2D- and 3D pelvic and leg kinematic strategies employed by the common quail to negotiate visible steps (upwards and downwards) of about 10%, and 50% of their leg length. We used biplanar fluoroscopy to accurately describe joint positions in three dimensions and performed semi-automatic landmark localization using deep learning. Quails negotiated the vertical obstacles without major problems and rapidly regained steady-state locomotion. When coping with step upwards, the quail mostly adapted the trailing limb to permit the leading leg to step on the elevated substrate similarly as it did during level locomotion. When negotiated steps downwards, both legs showed significant adaptations. For those small and moderate step heights that did not induce aerial running, the quail kept the kinematic pattern of the distal joints largely unchanged during uneven locomotion, and most changes occurred in proximal joints. The hip regulated leg length, while the distal joints maintained the spring-damped limb patterns. However, to negotiate the largest visible steps, more dramatic kinematic alterations were observed. There all joints contributed to leg lengthening/shortening in the trailing leg, and both the trailing and leading legs stepped more vertically and less abducted. In addition, locomotion speed was decreased. We hypothesize a shift from a dynamic walking program to more goal-directed motions that might be focused on maximizing safety.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Adjustments of global and local hindlimb properties during terrestrial locomotion of the common quail (Coturnix coturnix).J Exp Biol. 2013 Oct 15;216(Pt 20):3906-16. doi: 10.1242/jeb.085399. Epub 2013 Jul 18. J Exp Biol. 2013. PMID: 23868846
-
Don't break a leg: running birds from quail to ostrich prioritise leg safety and economy on uneven terrain.J Exp Biol. 2014 Nov 1;217(Pt 21):3786-96. doi: 10.1242/jeb.102640. J Exp Biol. 2014. PMID: 25355848 Free PMC article.
-
Kinematic trajectories in response to speed perturbations in walking suggest modular task-level control of leg angle and length.Integr Comp Biol. 2022 May 24:icac057. doi: 10.1093/icb/icac057. Online ahead of print. Integr Comp Biol. 2022. PMID: 35612979
-
Scaling of avian bipedal locomotion reveals independent effects of body mass and leg posture on gait.J Exp Biol. 2018 May 22;221(Pt 10):jeb152538. doi: 10.1242/jeb.152538. J Exp Biol. 2018. PMID: 29789347 Review.
-
Contributions to the understanding of gait control.Dan Med J. 2014 Apr;61(4):B4823. Dan Med J. 2014. PMID: 24814597 Review.
References
-
- Kilbourne BM, Andrada E, Fischer MS, Nyakatura JA. Morphology and motion: Hindlimb proportions and swing phase kinematics in terrestrially locomoting charadriiform birds. J. Exp. Biol. 2016;219:1405–1416. - PubMed
-
- Andrada E, Nyakatura JA, Bergmann F, Blickhan R. Adjustments of global and local hindlimb properties during terrestrial locomotion of the common quail (Coturnix coturnix) J. Exp. Biol. 2013;216:3906–3916. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
