

Interferon-Inducible E3 Ligase RNF213 Facilitates Host-Protective Linear and K63-Linked Ubiquitylation of Toxoplasma gondii Parasitophorous Vacuoles
- PMID: 36154443
- PMCID: PMC9601232
- DOI: 10.1128/mbio.01888-22
Interferon-Inducible E3 Ligase RNF213 Facilitates Host-Protective Linear and K63-Linked Ubiquitylation of Toxoplasma gondii Parasitophorous Vacuoles
Abstract
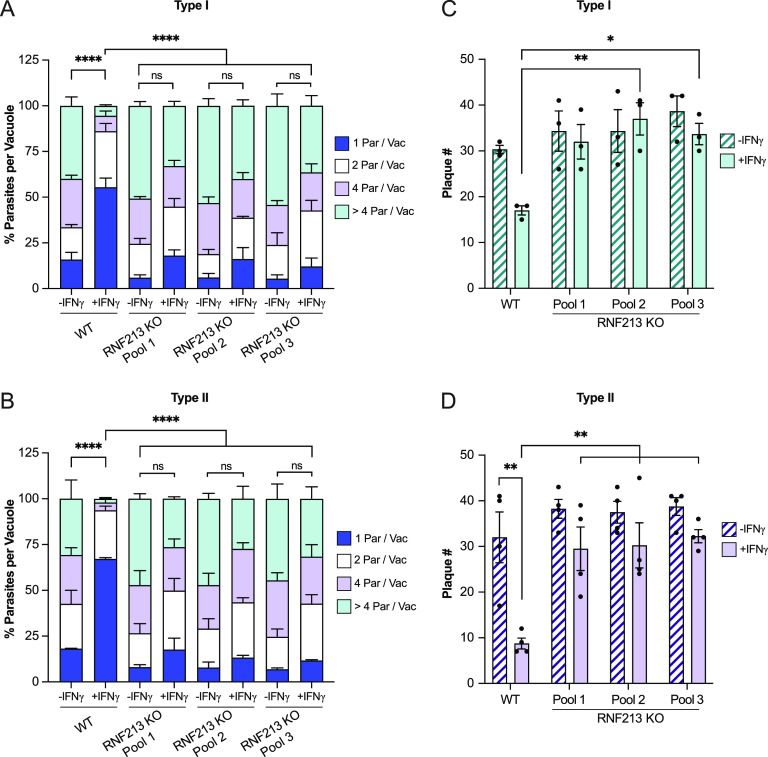
The obligate intracellular protozoan pathogen Toxoplasma gondii infects a wide range of vertebrate hosts and frequently causes zoonotic infections in humans. Whereas infected immunocompetent individuals typically remain asymptomatic, toxoplasmosis in immunocompromised individuals can manifest as a severe, potentially lethal disease, and congenital Toxoplasma infections are associated with adverse pregnancy outcomes. The protective immune response of healthy individuals involves the production of lymphocyte-derived cytokines such as interferon gamma (IFN-γ), which elicits cell-autonomous immunity in host cells. IFN-γ-inducible antiparasitic defense programs comprise nutritional immunity, the production of noxious gases, and the ubiquitylation of the Toxoplasma-containing parasitophorous vacuole (PV). PV ubiquitylation prompts the recruitment of host defense proteins to the PV and the consequential execution of antimicrobial effector programs, which reduce parasitic burden. However, the ubiquitin E3 ligase orchestrating these events has remained unknown. Here, we demonstrate that the IFN-γ-inducible E3 ligase RNF213 translocates to Toxoplasma PVs and facilitates PV ubiquitylation in human cells. Toxoplasma PVs become decorated with linear and K63-linked ubiquitin and recruit ubiquitin adaptor proteins in a process that is RNF213 dependent but independent of the linear ubiquitin chain assembly complex (LUBAC). IFN-γ priming fails to restrict Toxoplasma growth in cells lacking RNF213 expression, thus identifying RNF213 as a potent executioner of ubiquitylation-driven antiparasitic host defense. IMPORTANCE Globally, approximately one out of three people become infected with the obligate intracellular parasite Toxoplasma. These infections are typically asymptomatic but can cause severe disease and mortality in immunocompromised individuals. Infections can also be passed on from mother to fetus during pregnancy, potentially causing miscarriage or stillbirth. Therefore, toxoplasmosis constitutes a substantial public health burden. A better understanding of mechanisms by which healthy individuals control Toxoplasma infections could provide roadmaps toward novel therapies for vulnerable groups. Our work reveals a fundamental mechanism controlling intracellular Toxoplasma infections. Cytokines produced during Toxoplasma infections instruct human cells to produce the enzyme RNF213. We find that RNF213 labels intracellular vacuoles containing Toxoplasma with the small protein ubiquitin, which functions as an "eat-me" signal, attracting antimicrobial defense programs to fight off infection. Our work therefore identified a novel antiparasitic protein orchestrating a central aspect of the human immune response to Toxoplasma.
Keywords: LUBAC; RNF213; TAX1BP1; Toxoplasma; cell-autonomous immunity; interferons; linear ubiquitin; mysterin; parasitology; parasitophorous vacuole; ubiquitination; xenophagy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
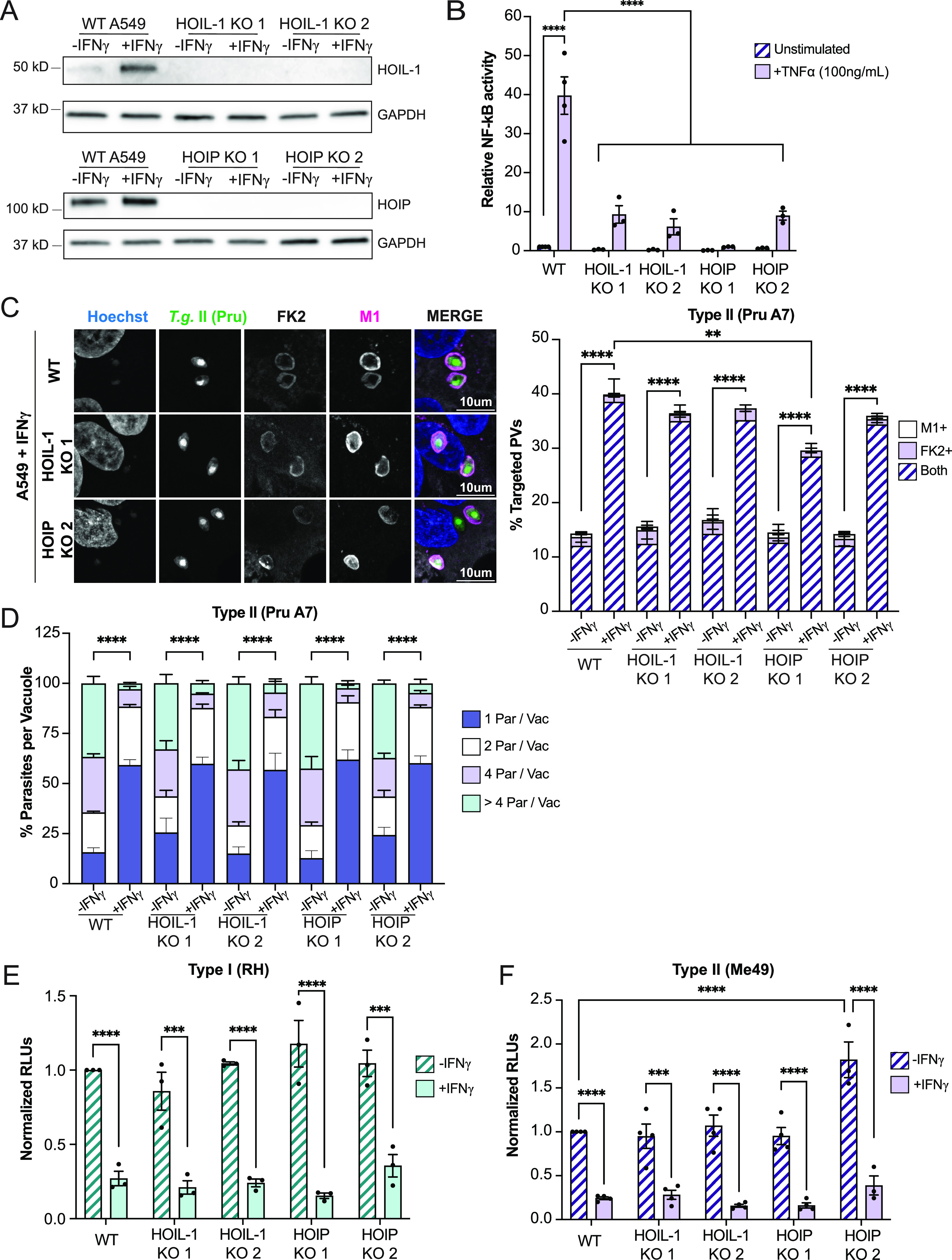
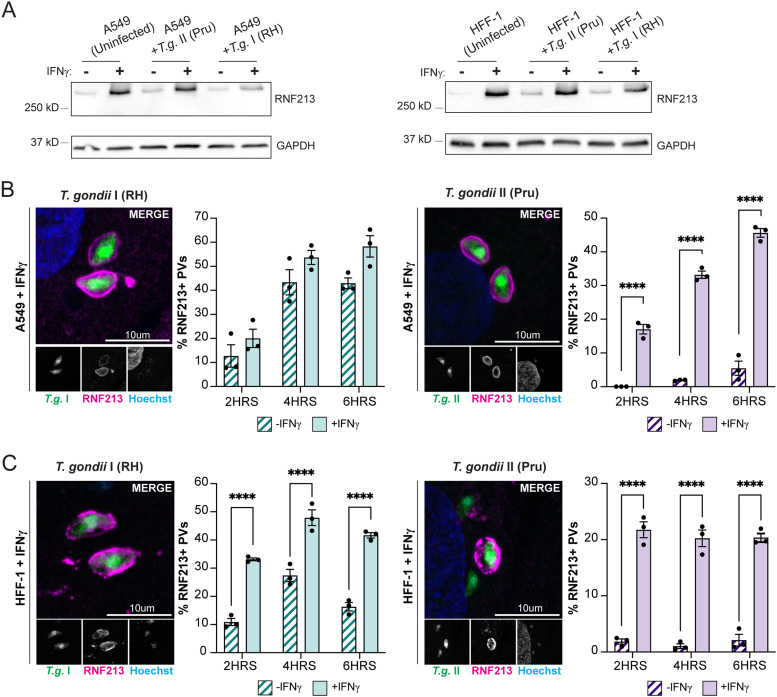
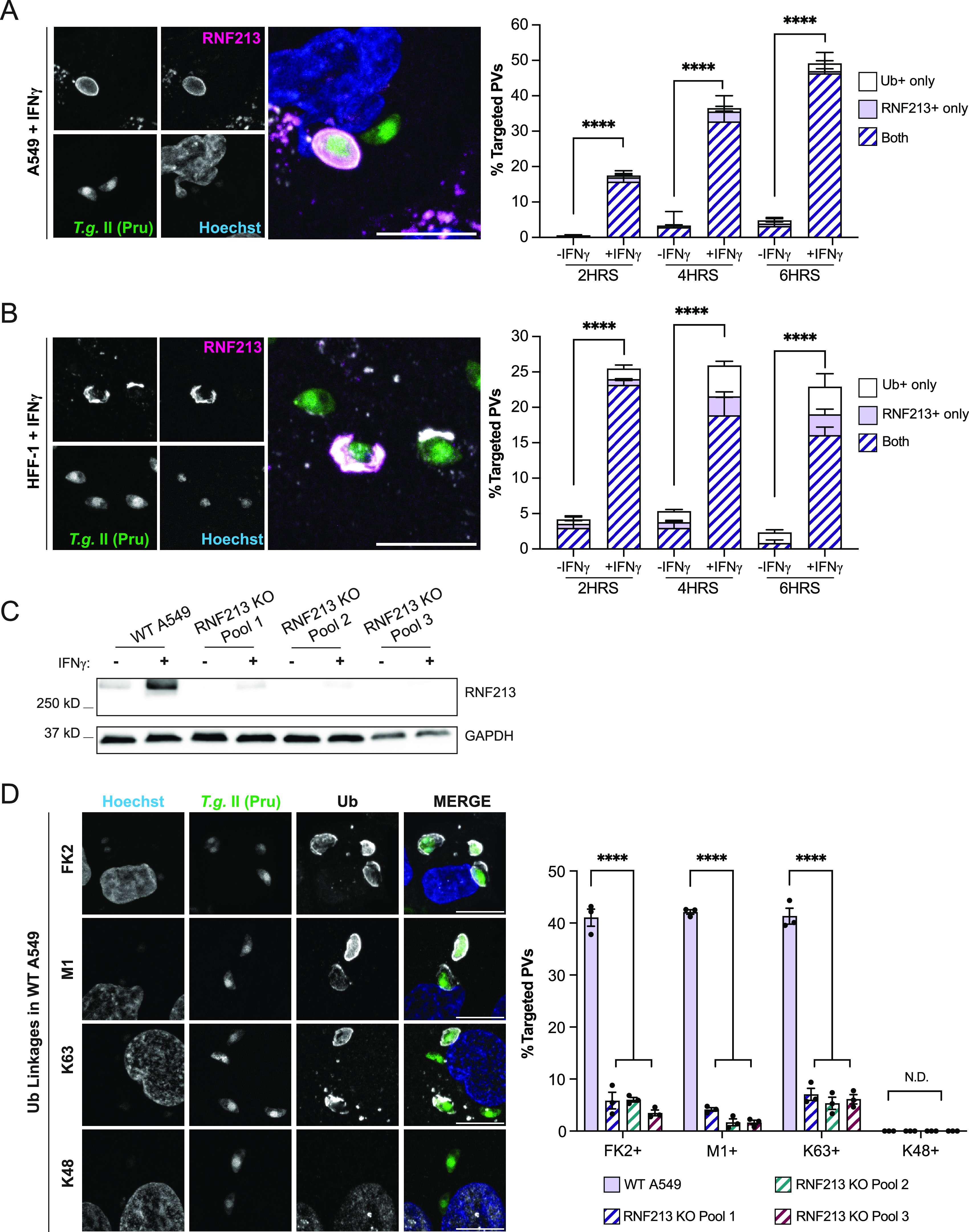
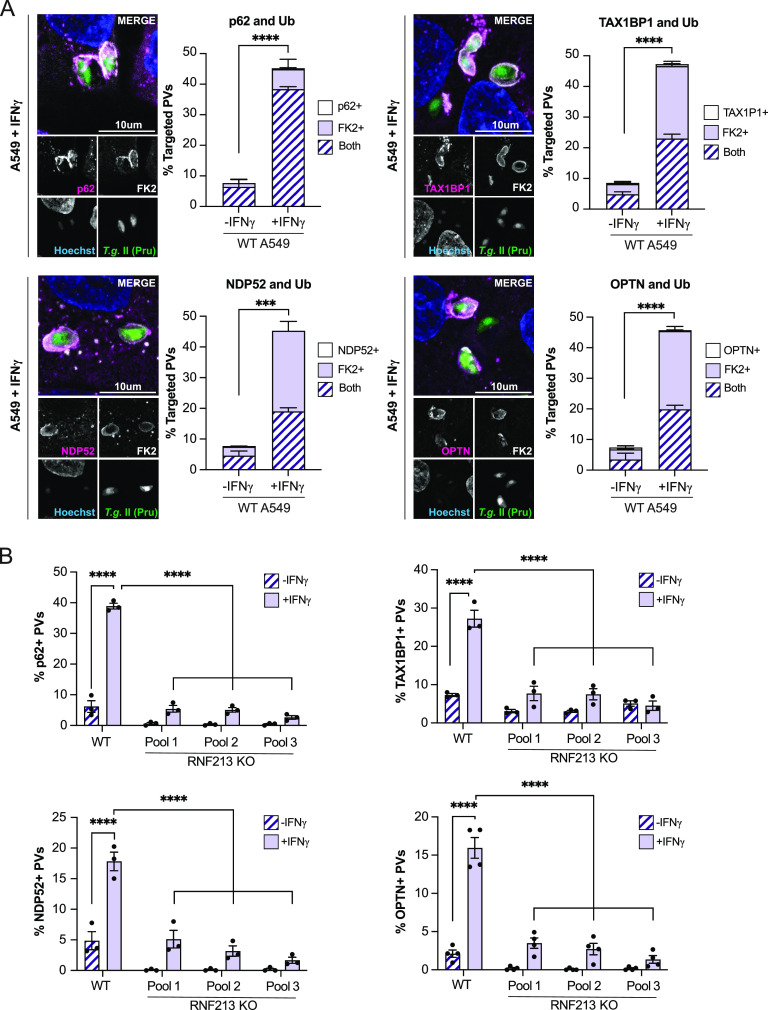
Similar articles
-
The emerging role of E3 ubiquitin ligase RNF213 as an antimicrobial host determinant.Front Cell Infect Microbiol. 2023 Aug 15;13:1205355. doi: 10.3389/fcimb.2023.1205355. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37655297 Free PMC article. Review.
-
Genome-wide and targeted CRISPR screens identify RNF213 as a mediator of interferon gamma-dependent pathogen restriction in human cells.Proc Natl Acad Sci U S A. 2024 Jan 2;121(1):e2315865120. doi: 10.1073/pnas.2315865120. Epub 2023 Dec 26. Proc Natl Acad Sci U S A. 2024. PMID: 38147552 Free PMC article.
-
Host E3 ubiquitin ligase ITCH mediates Toxoplasma gondii effector GRA35-triggered NLRP1 inflammasome activation and cell-autonomous immunity.mBio. 2024 Mar 13;15(3):e0330223. doi: 10.1128/mbio.03302-23. Epub 2024 Feb 20. mBio. 2024. PMID: 38376248 Free PMC article.
-
Murine Irgm Paralogs Regulate Nonredundant Functions To Execute Host Defense to Toxoplasma gondii.Infect Immun. 2021 Oct 15;89(11):e0020221. doi: 10.1128/IAI.00202-21. Epub 2021 Aug 2. Infect Immun. 2021. PMID: 34338548 Free PMC article.
-
The role of IFN-γ-mediated host immune responses in monitoring and the elimination of Toxoplasma gondii infection.Int Immunol. 2024 Apr 3;36(5):199-210. doi: 10.1093/intimm/dxae001. Int Immunol. 2024. PMID: 38175650 Free PMC article. Review.
Cited by
-
Insights into non-proteinaceous ubiquitination.Biochem Soc Trans. 2025 Apr 3;53(2):399-407. doi: 10.1042/BST20253029. Biochem Soc Trans. 2025. PMID: 40181599 Free PMC article. Review.
-
Host E3 ubiquitin ligase ITCH mediates Toxoplasma gondii effector GRA35-triggered NLRP1 inflammasome activation and cell-autonomous immunity.bioRxiv [Preprint]. 2023 Dec 14:2023.12.13.571530. doi: 10.1101/2023.12.13.571530. bioRxiv. 2023. Update in: mBio. 2024 Mar 13;15(3):e0330223. doi: 10.1128/mbio.03302-23. PMID: 38168400 Free PMC article. Updated. Preprint.
-
CRISPR Screens Identify Toxoplasma Genes That Determine Parasite Fitness in Interferon Gamma-Stimulated Human Cells.mBio. 2023 Apr 25;14(2):e0006023. doi: 10.1128/mbio.00060-23. Epub 2023 Mar 14. mBio. 2023. PMID: 36916910 Free PMC article.
-
The emerging role of E3 ubiquitin ligase RNF213 as an antimicrobial host determinant.Front Cell Infect Microbiol. 2023 Aug 15;13:1205355. doi: 10.3389/fcimb.2023.1205355. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37655297 Free PMC article. Review.
-
Bacterial esterases reverse lipopolysaccharide ubiquitylation to block host immunity.Cell Host Microbe. 2024 Jun 12;32(6):913-924.e7. doi: 10.1016/j.chom.2024.04.012. Cell Host Microbe. 2024. PMID: 38870903 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
