

Egg-adaptive mutations of human influenza H3N2 virus are contingent on natural evolution
- PMID: 36155668
- PMCID: PMC9536752
- DOI: 10.1371/journal.ppat.1010875
Egg-adaptive mutations of human influenza H3N2 virus are contingent on natural evolution
Abstract
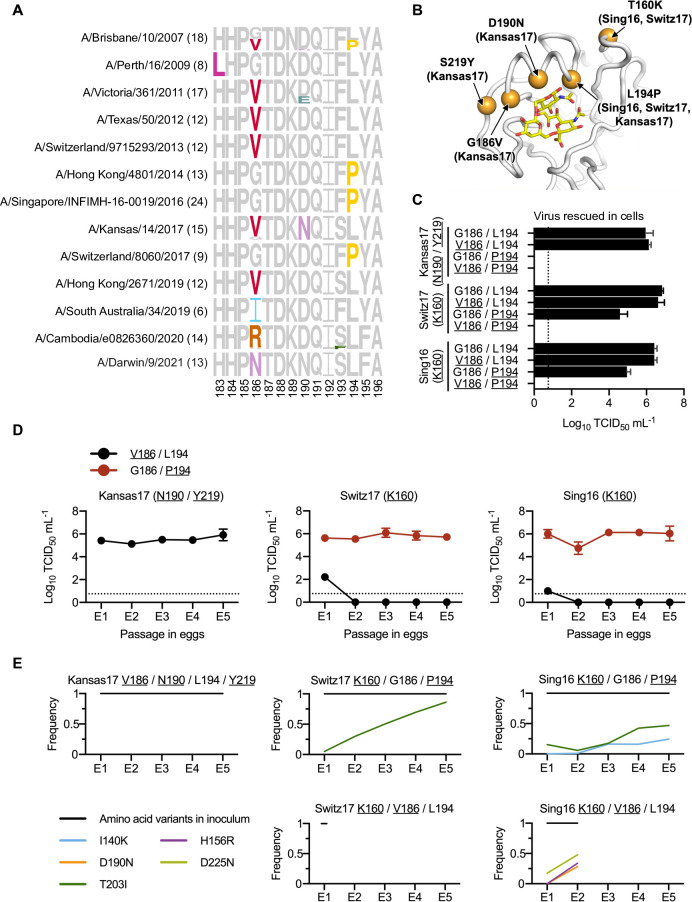
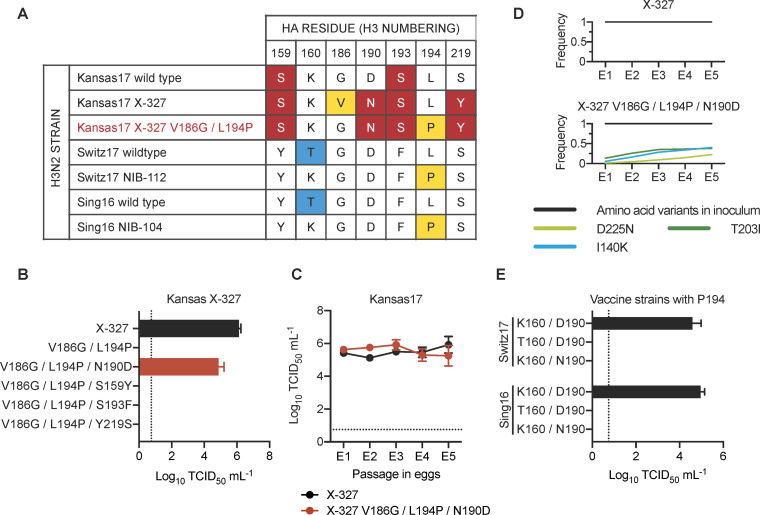
Egg-adaptive mutations in influenza hemagglutinin (HA) often emerge during the production of egg-based seasonal influenza vaccines, which contribute to the largest share in the global influenza vaccine market. While some egg-adaptive mutations have minimal impact on the HA antigenicity (e.g. G186V), others can alter it (e.g. L194P). Here, we show that the preference of egg-adaptive mutation in human H3N2 HA is strain-dependent. In particular, Thr160 and Asn190, which are found in many recent H3N2 strains, restrict the emergence of L194P but not G186V. Our results further suggest that natural amino acid variants at other HA residues also play a role in determining the preference of egg-adaptive mutation. Consistently, recent human H3N2 strains from different clades acquire different mutations during egg passaging. Overall, these results demonstrate that natural mutations in human H3N2 HA can influence the preference of egg-adaptation mutation, which has important implications in seed strain selection for egg-based influenza vaccine.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Preventing an Antigenically Disruptive Mutation in Egg-Based H3N2 Seasonal Influenza Vaccines by Mutational Incompatibility.Cell Host Microbe. 2019 Jun 12;25(6):836-844.e5. doi: 10.1016/j.chom.2019.04.013. Epub 2019 May 28. Cell Host Microbe. 2019. PMID: 31151913 Free PMC article.
-
Comparison of A(H3N2) Neutralizing Antibody Responses Elicited by 2018-2019 Season Quadrivalent Influenza Vaccines Derived from Eggs, Cells, and Recombinant Hemagglutinin.Clin Infect Dis. 2021 Dec 6;73(11):e4312-e4320. doi: 10.1093/cid/ciaa1352. Clin Infect Dis. 2021. PMID: 32898271 Clinical Trial.
-
In-depth phylogenetic analysis of the hemagglutinin gene of influenza A(H3N2) viruses circulating during the 2016-2017 season revealed egg-adaptive mutations of vaccine strains.Expert Rev Vaccines. 2020 Jan;19(1):115-122. doi: 10.1080/14760584.2020.1709827. Epub 2020 Jan 19. Expert Rev Vaccines. 2020. PMID: 31875483
-
Nucleoside-Modified mRNA-Based Influenza Vaccines Circumvent Problems Associated with H3N2 Vaccine Strain Egg Adaptation.J Virol. 2023 Jan 31;97(1):e0172322. doi: 10.1128/jvi.01723-22. Epub 2022 Dec 19. J Virol. 2023. PMID: 36533954 Free PMC article.
-
H3N2 influenza viruses in humans: Viral mechanisms, evolution, and evaluation.Hum Vaccin Immunother. 2018;14(8):1840-1847. doi: 10.1080/21645515.2018.1462639. Epub 2018 May 14. Hum Vaccin Immunother. 2018. PMID: 29641358 Free PMC article. Review.
Cited by
-
Predictive evolutionary modelling for influenza virus by site-based dynamics of mutations.Nat Commun. 2024 Mar 21;15(1):2546. doi: 10.1038/s41467-024-46918-0. Nat Commun. 2024. PMID: 38514647 Free PMC article.
-
Recent advances in the influenza virus vaccine landscape: a comprehensive overview of technologies and trials.Clin Microbiol Rev. 2024 Dec 10;37(4):e0002524. doi: 10.1128/cmr.00025-24. Epub 2024 Oct 3. Clin Microbiol Rev. 2024. PMID: 39360831 Review.
-
Adaptive truncation of the S gene in IBV during chicken embryo passaging plays a crucial role in its attenuation.PLoS Pathog. 2024 Jul 30;20(7):e1012415. doi: 10.1371/journal.ppat.1012415. eCollection 2024 Jul. PLoS Pathog. 2024. PMID: 39078847 Free PMC article.
-
Seasonal antigenic prediction of influenza A H3N2 using machine learning.Nat Commun. 2024 May 7;15(1):3833. doi: 10.1038/s41467-024-47862-9. Nat Commun. 2024. PMID: 38714654 Free PMC article.
-
A Phase 1/2 Randomized Study to Evaluate the Safety, Tolerability, and Immunogenicity of Nucleoside-Modified Messenger RNA Influenza Vaccines in Healthy Adults.Vaccines (Basel). 2025 Apr 3;13(4):383. doi: 10.3390/vaccines13040383. Vaccines (Basel). 2025. PMID: 40333267 Free PMC article.
References
-
- Centers for Disease Control and Prevention. Past seasons vaccine effectiveness estimates [26 August, 2021]. Available from: https://www.cdc.gov/flu/vaccines-work/past-seasons-estimates.html.
-
- Belongia EA, Simpson MD, King JP, Sundaram ME, Kelley NS, Osterholm MT, et al.. Variable influenza vaccine effectiveness by subtype: a systematic review and meta-analysis of test-negative design studies. Lancet Infect Dis. 2016;16(8):942–51. Epub 20160406. doi: 10.1016/S1473-3099(16)00129-8 . - DOI - PubMed
-
- Okoli GN, Racovitan F, Abdulwahid T, Righolt CH, Mahmud SM. Variable seasonal influenza vaccine effectiveness across geographical regions, age groups and levels of vaccine antigenic similarity with circulating virus strains: a systematic review and meta-analysis of the evidence from test-negative design studies after the 2009/10 influenza pandemic. Vaccine. 2021;39(8):1225–40. Epub 20210122. doi: 10.1016/j.vaccine.2021.01.032 . - DOI - PubMed
-
- Kodihalli S, Justewicz DM, Gubareva LV, Webster RG. Selection of a single amino acid substitution in the hemagglutinin molecule by chicken eggs can render influenza A virus (H3) candidate vaccine ineffective. J Virol. 1995;69(8):4888–97. doi: 10.1128/JVI.69.8.4888-4897.1995 ; PubMed Central PMCID: PMC189303. - DOI - PMC - PubMed
-
- Zost SJ, Parkhouse K, Gumina ME, Kim K, Diaz Perez S, Wilson PC, et al.. Contemporary H3N2 influenza viruses have a glycosylation site that alters binding of antibodies elicited by egg-adapted vaccine strains. Proc Natl Acad Sci U S A. 2017;114(47):12578–83. Epub 2017/11/08. doi: 10.1073/pnas.1712377114 ; PubMed Central PMCID: PMC5703309. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
