

Detection of Homologous Recombination Intermediates via Proximity Ligation and Quantitative PCR in Saccharomyces cerevisiae
- PMID: 36155960
- PMCID: PMC10205173
- DOI: 10.3791/64240
Detection of Homologous Recombination Intermediates via Proximity Ligation and Quantitative PCR in Saccharomyces cerevisiae
Abstract
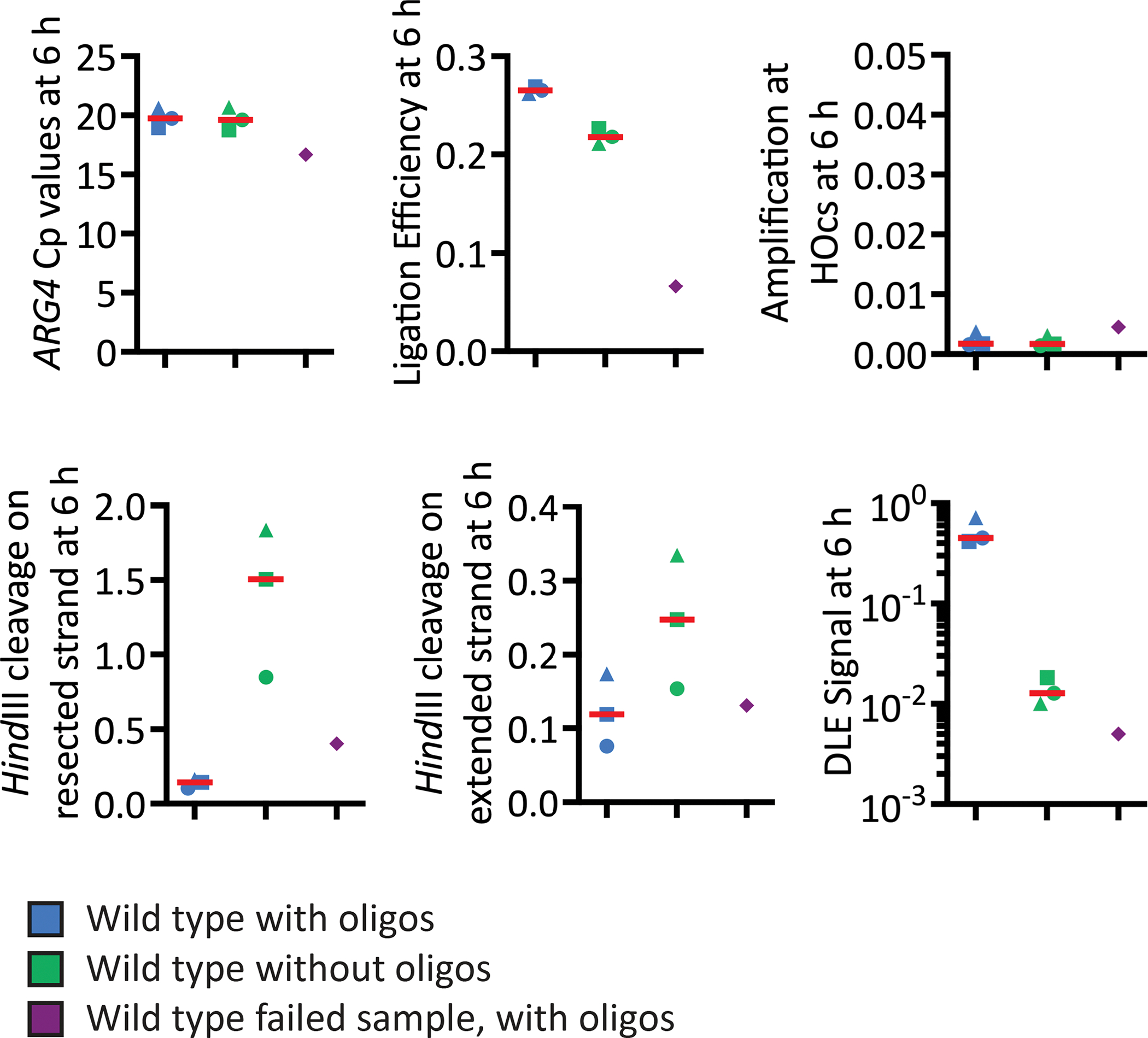
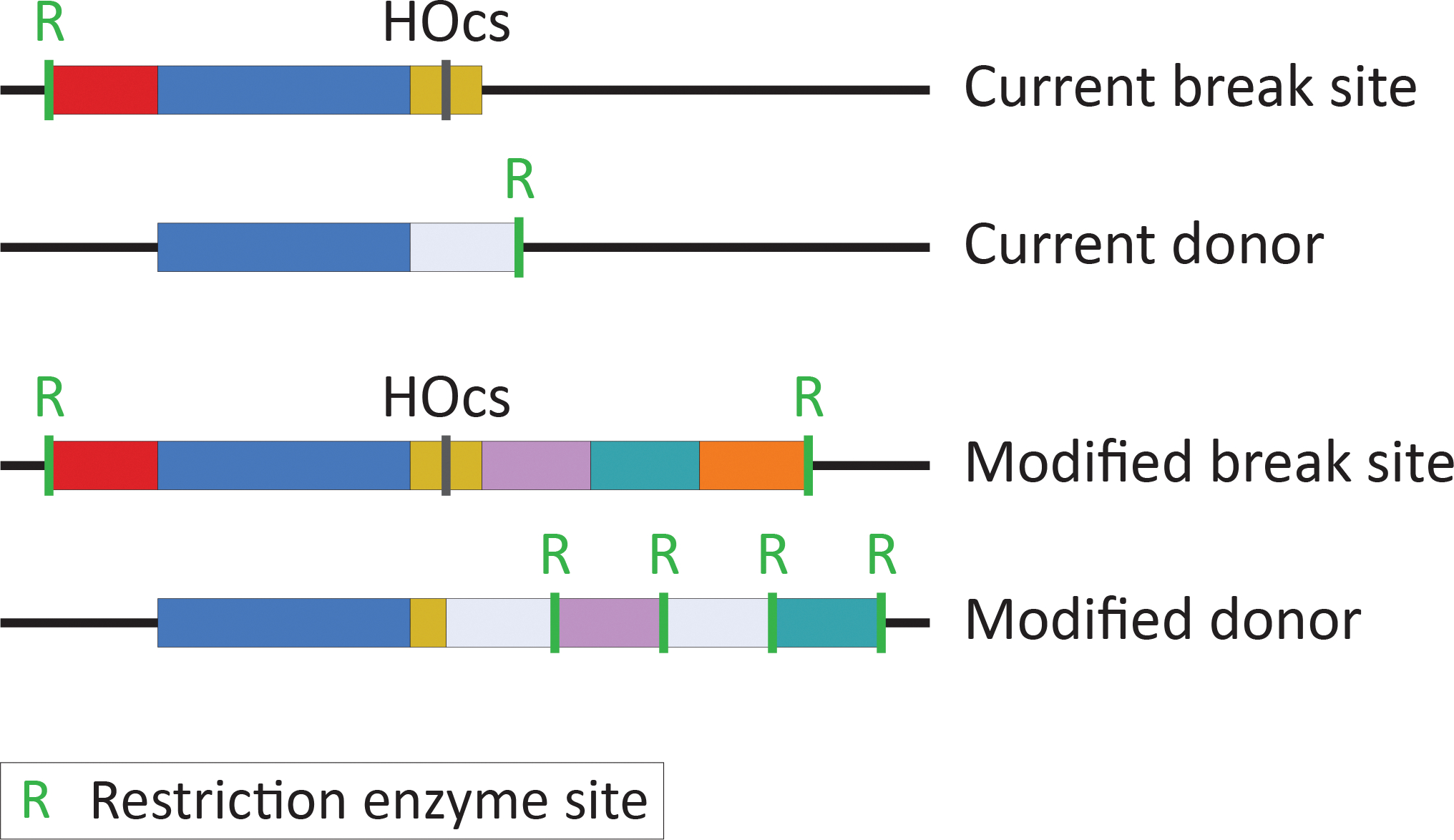
DNA damage, including DNA double-stranded breaks and inter-strand cross-links, incurred during the S and G2 phases of the cell cycle can be repaired by homologous recombination (HR). In addition, HR represents an important mechanism of replication fork rescue following stalling or collapse. The regulation of the many reversible and irreversible steps of this complex pathway promotes its fidelity. The physical analysis of the recombination intermediates formed during HR enables the characterization of these controls by various nucleoprotein factors and their interactors. Though there are well-established methods to assay specific events and intermediates in the recombination pathway, the detection of D-loop formation and extension, two critical steps in this pathway, has proved challenging until recently. Here, efficient methods for detecting key events in the HR pathway, namely DNA double-stranded break formation, D-loop formation, D-loop extension, and the formation of products via break-induced replication (BIR) in Saccharomyces cerevisiae are described. These assays detect their relevant recombination intermediates and products with high sensitivity and are independent of cellular viability. The detection of D-loops, D-loop extension, and the BIR product is based on proximity ligation. Together, these assays allow for the study of the kinetics of HR at the population level to finely address the functions of HR proteins and regulators at significant steps in the pathway.
Conflict of interest statement
DISCLOSURES:
The authors have nothing to disclose.
Figures




References
-
- Symington LS, Gautier J Double-strand break end resection and repair pathway choice. Annual Review of Genetics. 45 (1), 247–271 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
