

ER-mitochondria contact sites; a multifaceted factory for Ca2+ signaling and lipid transport
- PMID: 36158205
- PMCID: PMC9494157
- DOI: 10.3389/fcell.2022.988014
ER-mitochondria contact sites; a multifaceted factory for Ca2+ signaling and lipid transport
Abstract
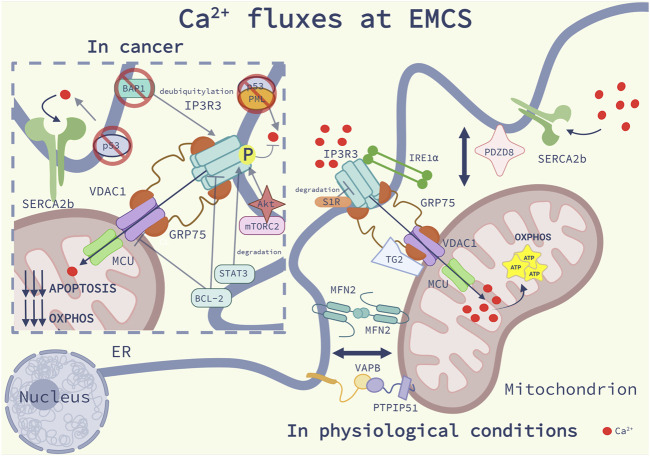
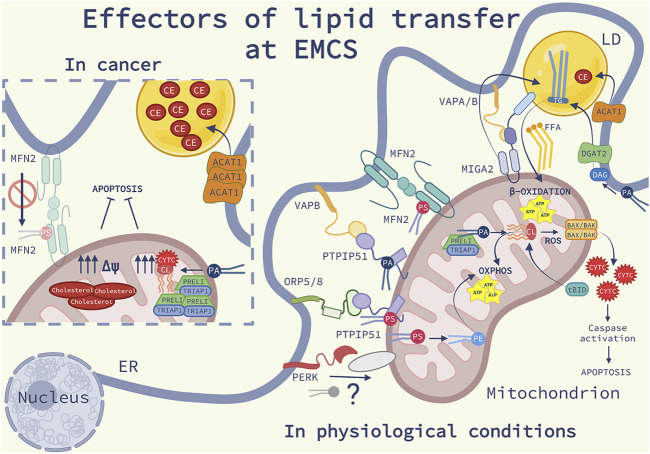
Membrane contact sites (MCS) between organelles of eukaryotic cells provide structural integrity and promote organelle homeostasis by facilitating intracellular signaling, exchange of ions, metabolites and lipids and membrane dynamics. Cataloguing MCS revolutionized our understanding of the structural organization of a eukaryotic cell, but the functional role of MSCs and their role in complex diseases, such as cancer, are only gradually emerging. In particular, the endoplasmic reticulum (ER)-mitochondria contacts (EMCS) are key effectors of non-vesicular lipid trafficking, thereby regulating the lipid composition of cellular membranes and organelles, their physiological functions and lipid-mediated signaling pathways both in physiological and diseased conditions. In this short review, we discuss key aspects of the functional complexity of EMCS in mammalian cells, with particular emphasis on their role as central hubs for lipid transport between these organelles and how perturbations of these pathways may favor key traits of cancer cells.
Keywords: Ca2+ signaling; ER-mitochondria contact sites; cancer; lipid transfer protein; lipids.
Copyright © 2022 Sassano, Felipe-Abrio and Agostinis.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous