

From Hair to Colon: Hair Follicle-Derived MSCs Alleviate Pyroptosis in DSS-Induced Ulcerative Colitis by Releasing Exosomes in a Paracrine Manner
- PMID: 36160717
- PMCID: PMC9507792
- DOI: 10.1155/2022/9097530
From Hair to Colon: Hair Follicle-Derived MSCs Alleviate Pyroptosis in DSS-Induced Ulcerative Colitis by Releasing Exosomes in a Paracrine Manner
Abstract
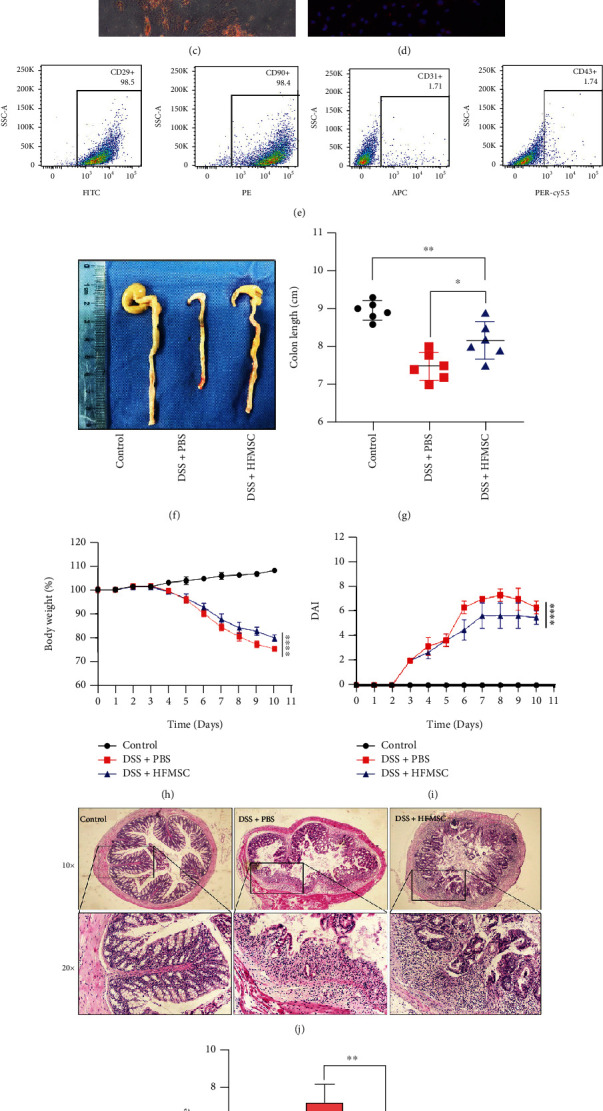
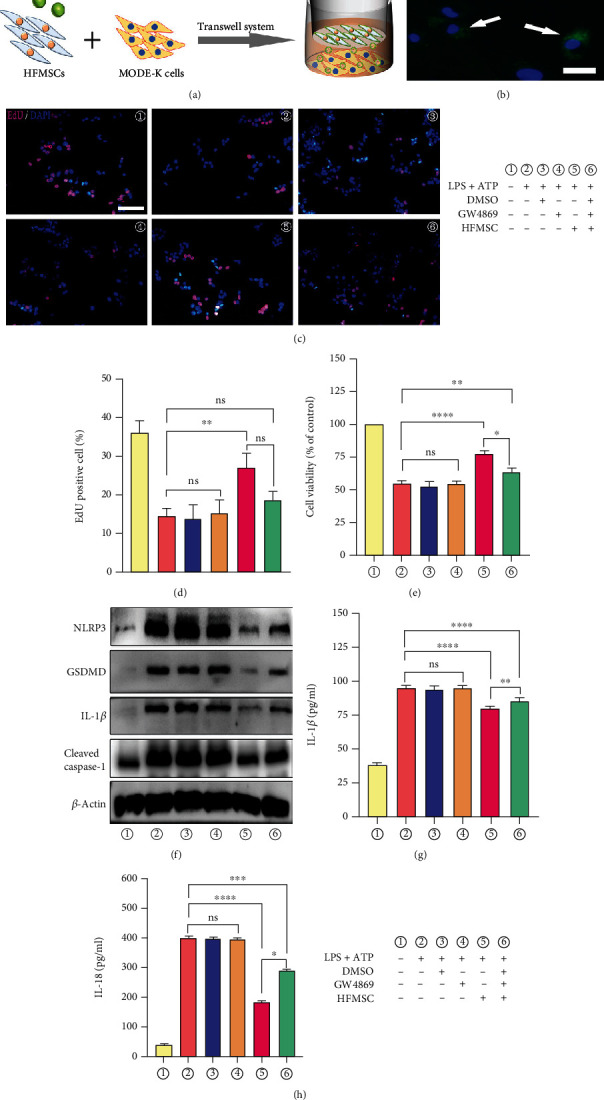
Ulcerative colitis (UC) has attracted intense attention due to its high recurrence rate and the difficulty of treatment. Pyroptosis has been suggested to be crucial in the development of UC. Although mesenchymal stem cells (MSCs) are broadly used for UC therapy, they have rarely been studied in the context of UC pyroptosis. Hair follicle-derived MSCs (HFMSCs) are especially understudied with regard to UC and pyroptosis. In this study, we aimed to discover the effects and potential mechanisms of HFMSCs in UC. We administered HFMSCs to dextran sulfate sodium- (DSS-) treated mice and found that the HFMSCs significantly inhibited pyroptosis to alleviate DSS-induced UC. A transwell system and GW4869, an exosome inhibitor, were used to prove the paracrine mechanism of HFMSCs. HFMSC supernatant reduced pyroptosis-related protein expression and promoted cell viability, but these effects were attenuated by GW4869, suggesting a role for HFMSC-released exosomes (Exos) in pyroptosis. Next, Exos were extracted and administered in vitro and in vivo to explore their roles in pyroptosis and UC. In addition, the biodistribution of Exos in mice was tracked using an imaging system and immunofluorescence. The results suggested that Exos not only improved DSS-induced pyroptosis and UC but also were internalized into the injured colon. Furthermore, the therapeutic efficacy of Exos was dose dependent. Among the Exo treatments, administration of 400 μg of Exos per mouse twice a week exhibited the highest efficacy. The differentially expressed miRNAs (DEmiRNAs) between MSCs and MSC-released Exos suggested that Exos might inhibit pyroptosis through tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) signalling and interferon- (IFN-) gamma pathways. Our study reveals that HFMSCs can alleviate pyroptosis in UC by releasing DEmiRNA-containing Exos in a paracrine manner. This finding may lead to new treatments for UC.
Copyright © 2022 Yuan Chang et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures





Similar articles
-
Stimulation by exosomes from hypoxia-preconditioned hair follicle mesenchymal stem cells facilitates mitophagy by inhibiting the PI3K/AKT/mTOR signaling pathway to alleviate ulcerative colitis.Theranostics. 2024 Jul 8;14(11):4278-4296. doi: 10.7150/thno.96038. eCollection 2024. Theranostics. 2024. PMID: 39113800 Free PMC article.
-
Exosomes Derived from Apelin-Pretreated Mesenchymal Stem Cells Ameliorate Sepsis-Induced Myocardial Dysfunction by Alleviating Cardiomyocyte Pyroptosis via Delivery of miR-34a-5p.Int J Nanomedicine. 2025 Jan 17;20:687-703. doi: 10.2147/IJN.S498770. eCollection 2025. Int J Nanomedicine. 2025. PMID: 39845770 Free PMC article.
-
Hair follicle mesenchymal stem cell exosomal lncRNA H19 inhibited NLRP3 pyroptosis to promote diabetic mouse skin wound healing.Aging (Albany NY). 2023 Feb 14;15(3):791-809. doi: 10.18632/aging.204513. Epub 2023 Feb 14. Aging (Albany NY). 2023. PMID: 36787444 Free PMC article.
-
The therapeutic potential of stem cell-derived exosomes in the ulcerative colitis and colorectal cancer.Stem Cell Res Ther. 2022 Apr 1;13(1):138. doi: 10.1186/s13287-022-02811-5. Stem Cell Res Ther. 2022. PMID: 35365226 Free PMC article. Review.
-
Recent advances in the delivery of microRNAs via exosomes derived from MSCs, and their role in regulation of ferroptosis.Pathol Res Pract. 2025 Jun;270:155984. doi: 10.1016/j.prp.2025.155984. Epub 2025 Apr 21. Pathol Res Pract. 2025. PMID: 40315562 Review.
Cited by
-
Mesenchymal stem cell-derived exosomes as a plausible immunomodulatory therapeutic tool for inflammatory diseases.Front Cell Dev Biol. 2025 Mar 10;13:1563427. doi: 10.3389/fcell.2025.1563427. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40129569 Free PMC article. Review.
-
Extracellular vesicles for the treatment of ulcerative colitis: A systematic review and meta-analysis of animal studies.Heliyon. 2024 Aug 24;10(17):e36890. doi: 10.1016/j.heliyon.2024.e36890. eCollection 2024 Sep 15. Heliyon. 2024. PMID: 39281542 Free PMC article.
-
Perinatal tissue-derived exosomes ameliorate colitis in mice by regulating the Foxp3 + Treg cells and gut microbiota.Stem Cell Res Ther. 2023 Mar 20;14(1):43. doi: 10.1186/s13287-023-03263-1. Stem Cell Res Ther. 2023. PMID: 36941715 Free PMC article.
-
Emerging role of exosomes in ulcerative colitis: Targeting NOD-like receptor family pyrin domain containing 3 inflammasome.World J Gastroenterol. 2024 Feb 14;30(6):527-541. doi: 10.3748/wjg.v30.i6.527. World J Gastroenterol. 2024. PMID: 38463022 Free PMC article. Review.
-
From stem cells to extracellular vesicles: a new horizon in tissue engineering and regenerative medicine.Cytotechnology. 2024 Aug;76(4):363-401. doi: 10.1007/s10616-024-00631-4. Epub 2024 May 8. Cytotechnology. 2024. PMID: 38933869 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
