

Structural view of G protein-coupled receptor signaling in the retinal rod outer segment
- PMID: 36163145
- PMCID: PMC9868064
- DOI: 10.1016/j.tibs.2022.08.010
Structural view of G protein-coupled receptor signaling in the retinal rod outer segment
Abstract
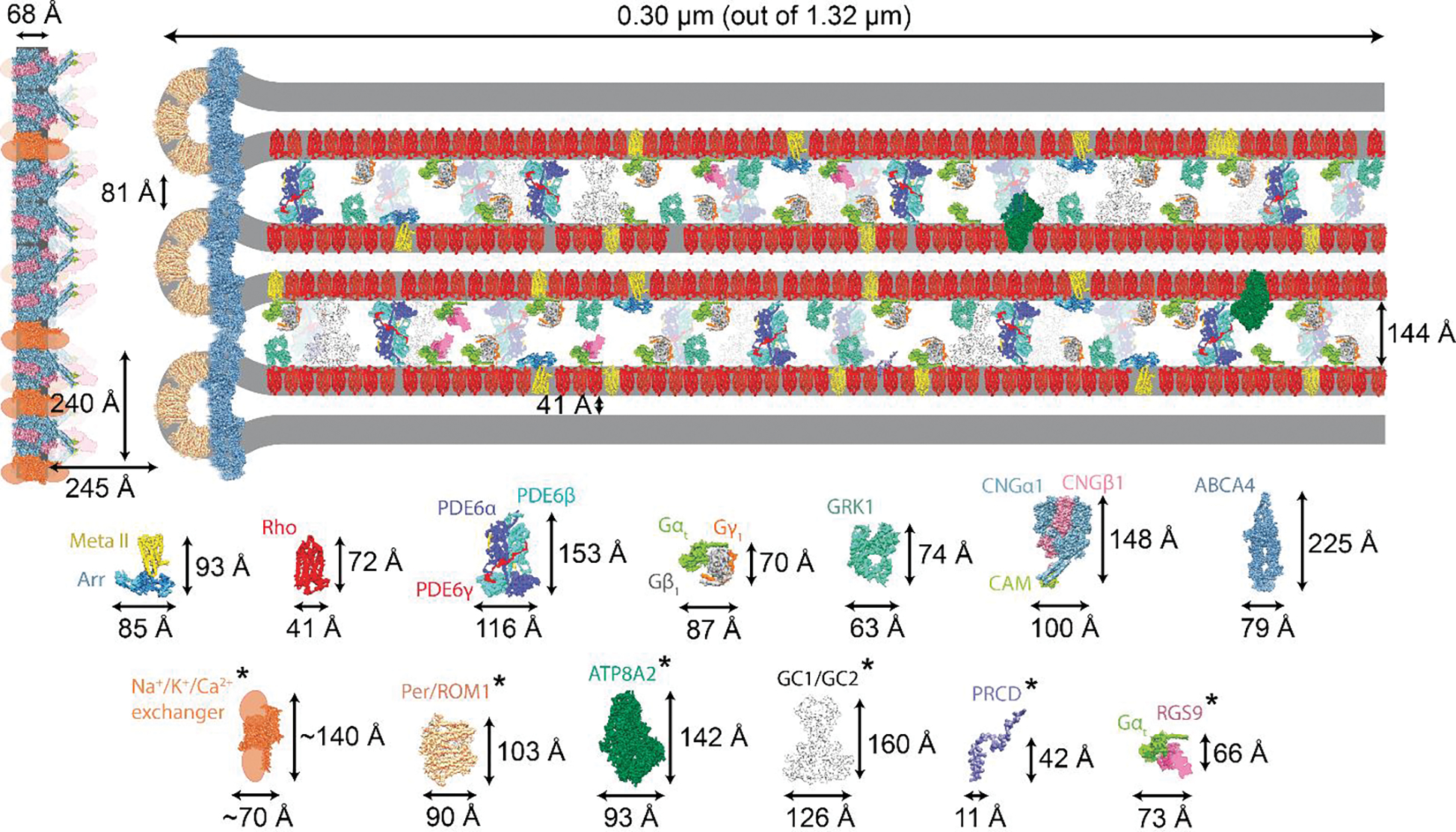
Visual phototransduction is the most extensively studied G protein-coupled receptor (GPCR) signaling pathway because of its quantifiable stimulus, non-redundancy of genes, and immense importance in vision. We summarize recent discoveries that have advanced our understanding of rod outer segment (ROS) morphology and the pathological basis of retinal diseases. We have combined recently published cryo-electron tomography (cryo-ET) data on the ROS with structural knowledge on individual proteins to define the precise spatial limitations under which phototransduction occurs. Although hypothetical, the reconstruction of the rod phototransduction system highlights the potential roles of phosphodiesterase 6 (PDE6) and guanylate cyclases (GCs) in maintaining the spacing between ROS discs, suggesting a plausible mechanism by which intrinsic optical signals are generated in the retina.
Keywords: G protein-coupled receptor (GPCR); GPCR signaling; phototransduction; rhodopsin; rod cell; rod outer segment (ROS).
Copyright © 2022 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest.
Figures




References
-
- Rodieck RW (1998) The first steps in seeing, Sinauer Associates.
-
- Polans A et al. (1996) Turned on by Ca2+! The physiology and pathology of Ca(2+)-binding proteins in the retina. Trends Neurosci 19 (12), 547–54. - PubMed
-
- Kawamura S and Tachibanaki S (2021) Molecular bases of rod and cone differences. Prog Retin Eye Res, 101040. - PubMed
