

Computational identification of a systemic antibiotic for gram-negative bacteria
- PMID: 36163500
- PMCID: PMC10155127
- DOI: 10.1038/s41564-022-01227-4
Computational identification of a systemic antibiotic for gram-negative bacteria
Abstract
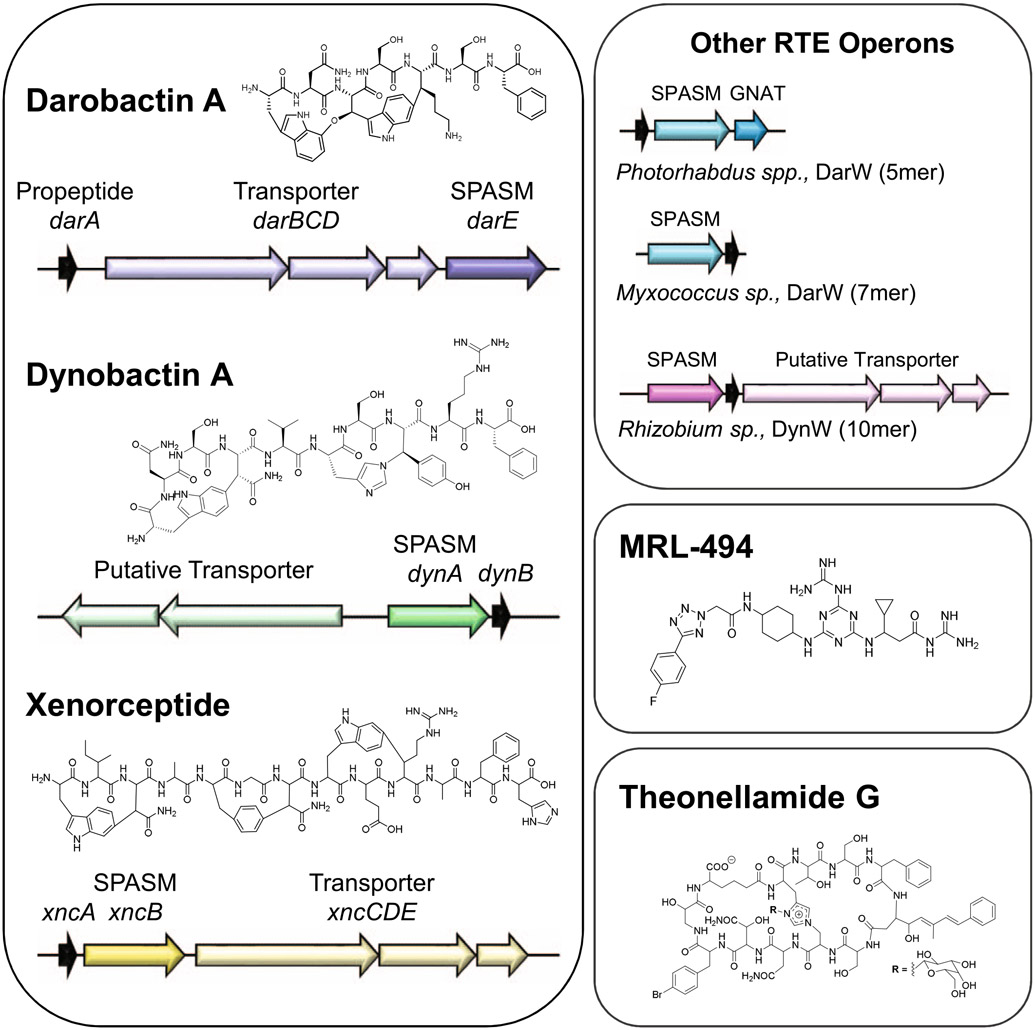
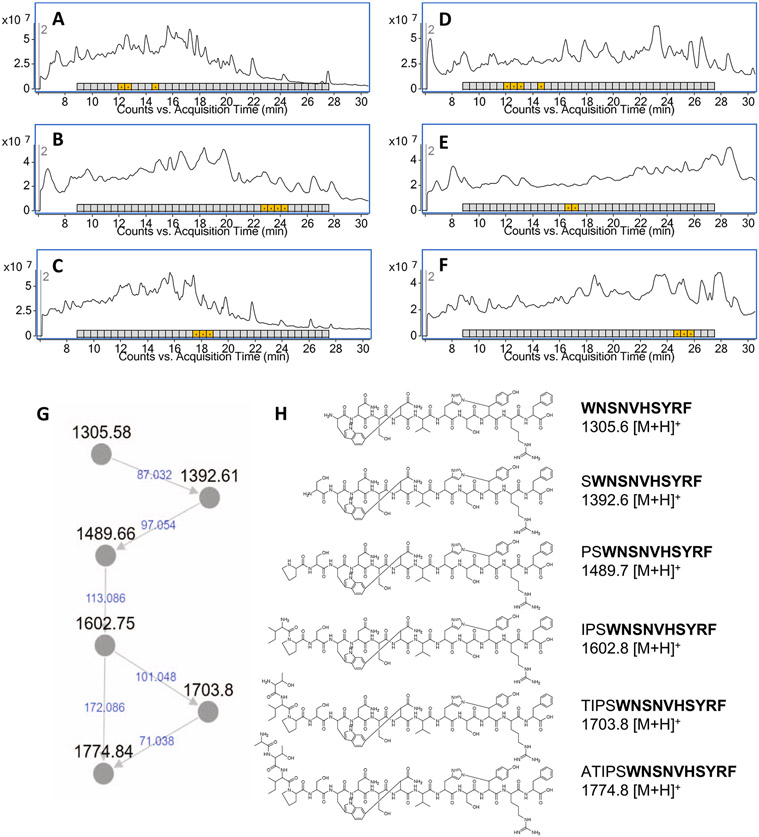
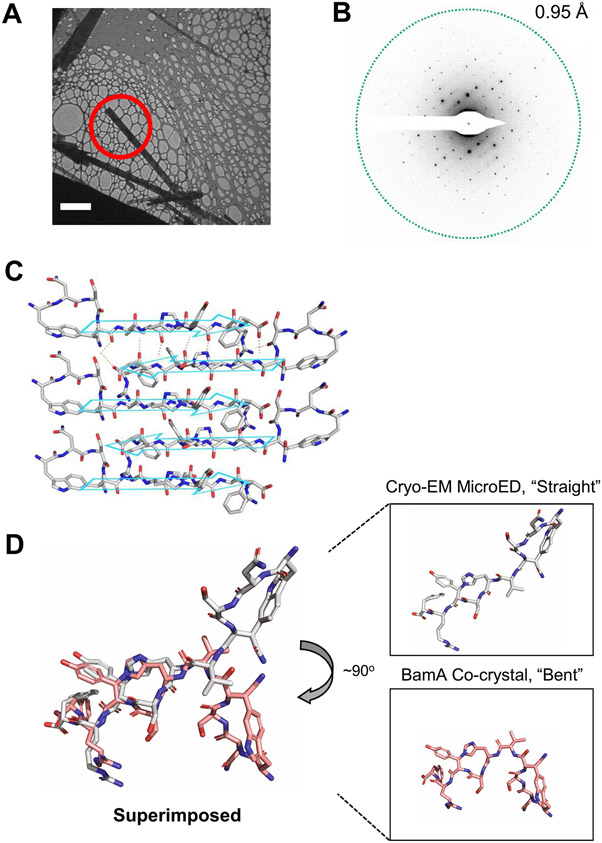
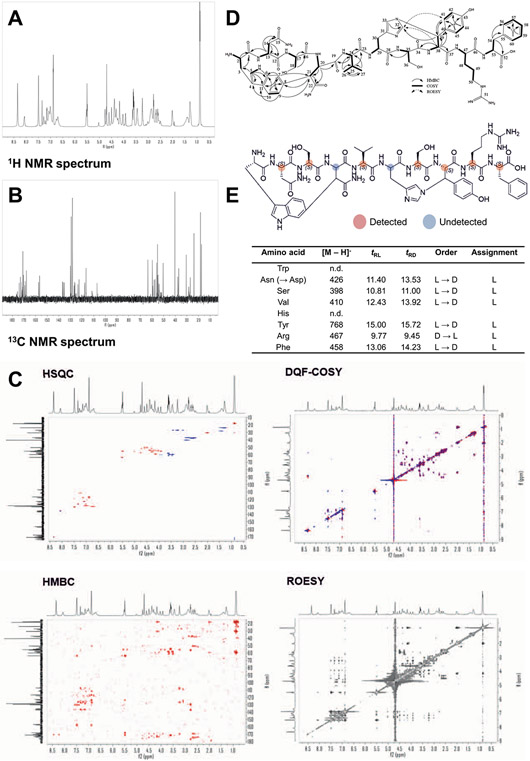
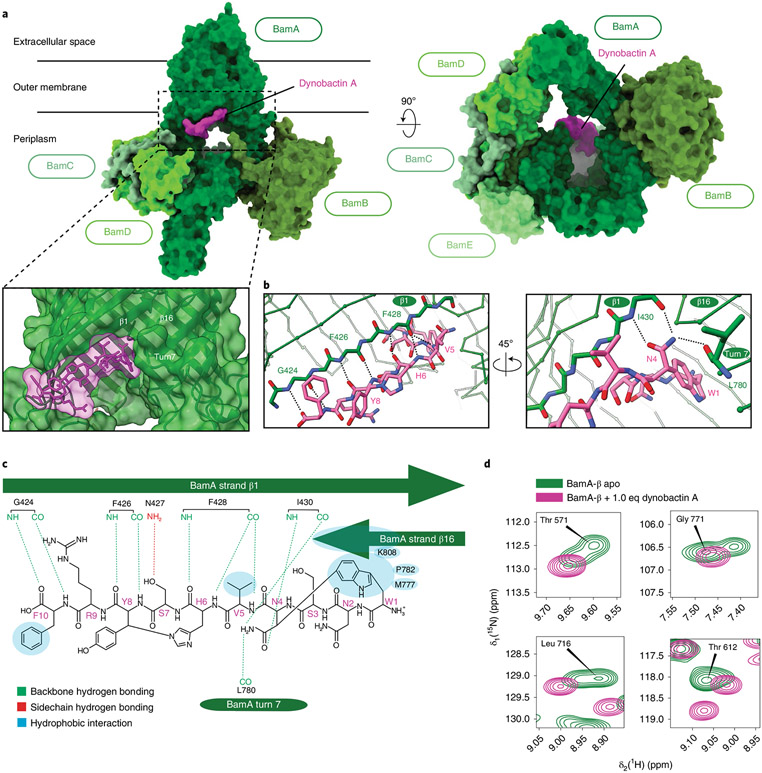
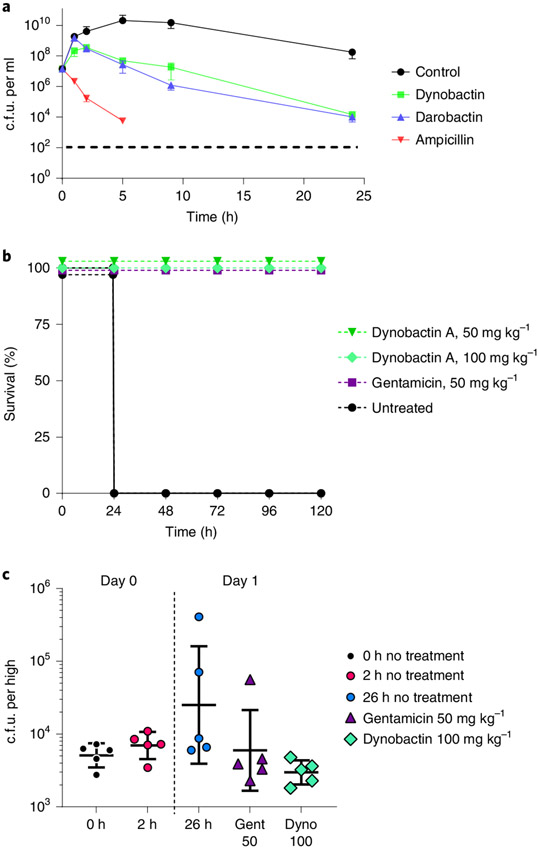
Discovery of antibiotics acting against Gram-negative species is uniquely challenging due to their restrictive penetration barrier. BamA, which inserts proteins into the outer membrane, is an attractive target due to its surface location. Darobactins produced by Photorhabdus, a nematode gut microbiome symbiont, target BamA. We reasoned that a computational search for genes only distantly related to the darobactin operon may lead to novel compounds. Following this clue, we identified dynobactin A, a novel peptide antibiotic from Photorhabdus australis containing two unlinked rings. Dynobactin is structurally unrelated to darobactins, but also targets BamA. Based on a BamA-dynobactin co-crystal structure and a BAM-complex-dynobactin cryo-EM structure, we show that dynobactin binds to the BamA lateral gate, uniquely protruding into its β-barrel lumen. Dynobactin showed efficacy in a mouse systemic Escherichia coli infection. This study demonstrates the utility of computational approaches to antibiotic discovery and suggests that dynobactin is a promising lead for drug development.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
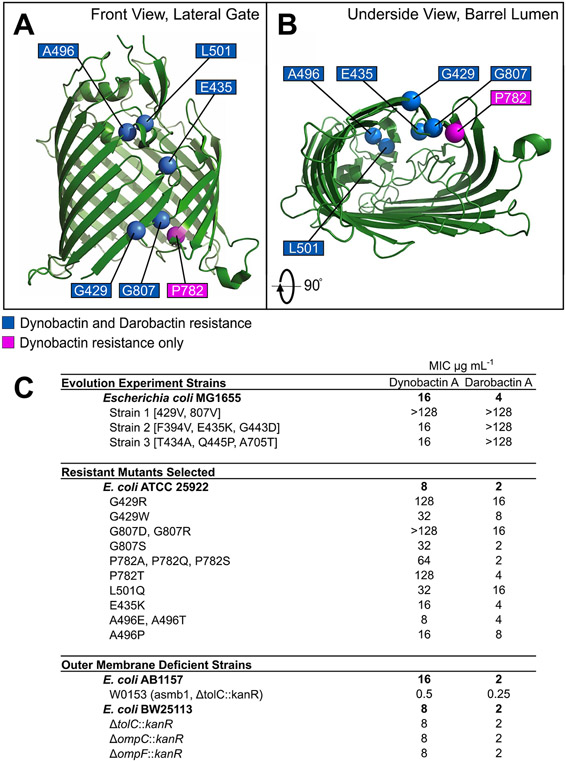
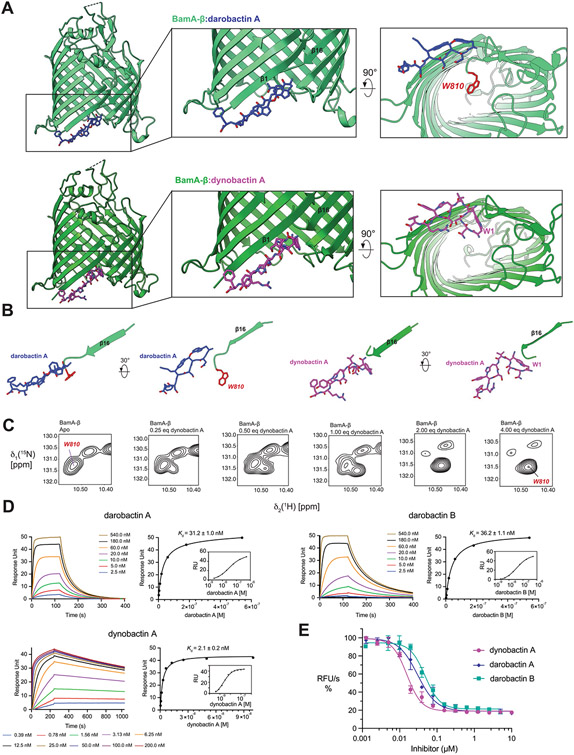
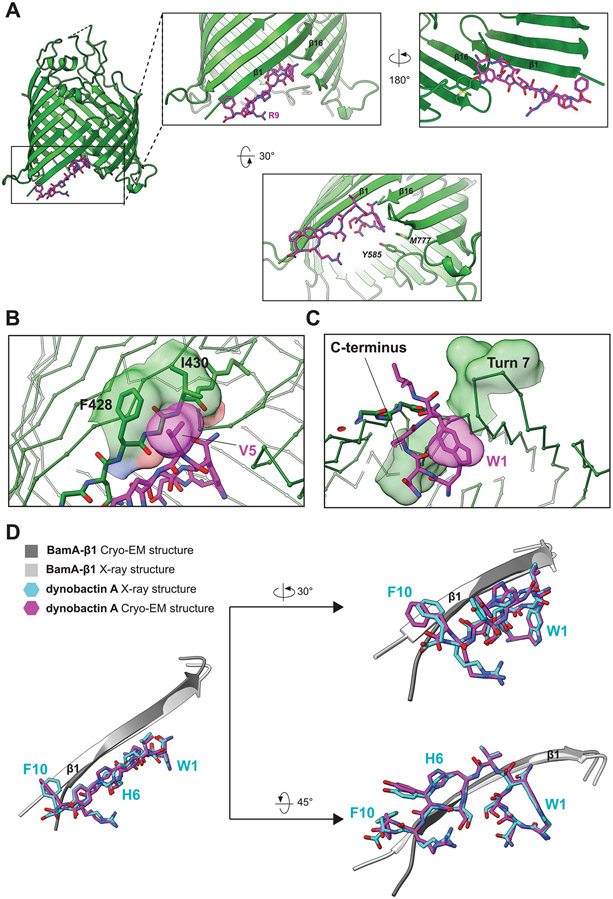
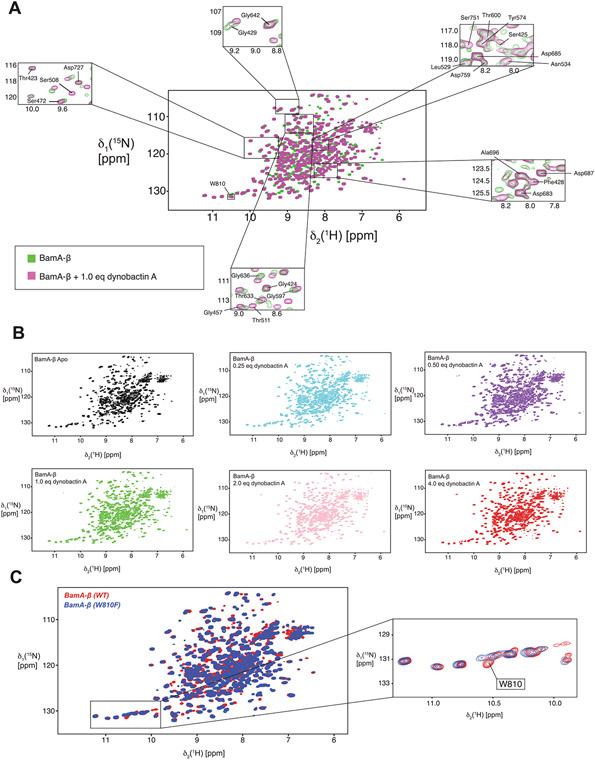
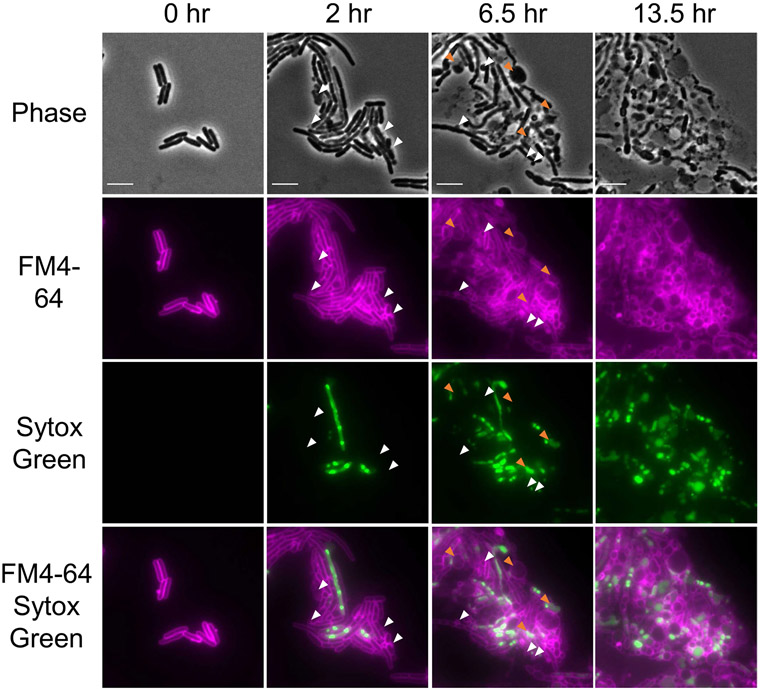
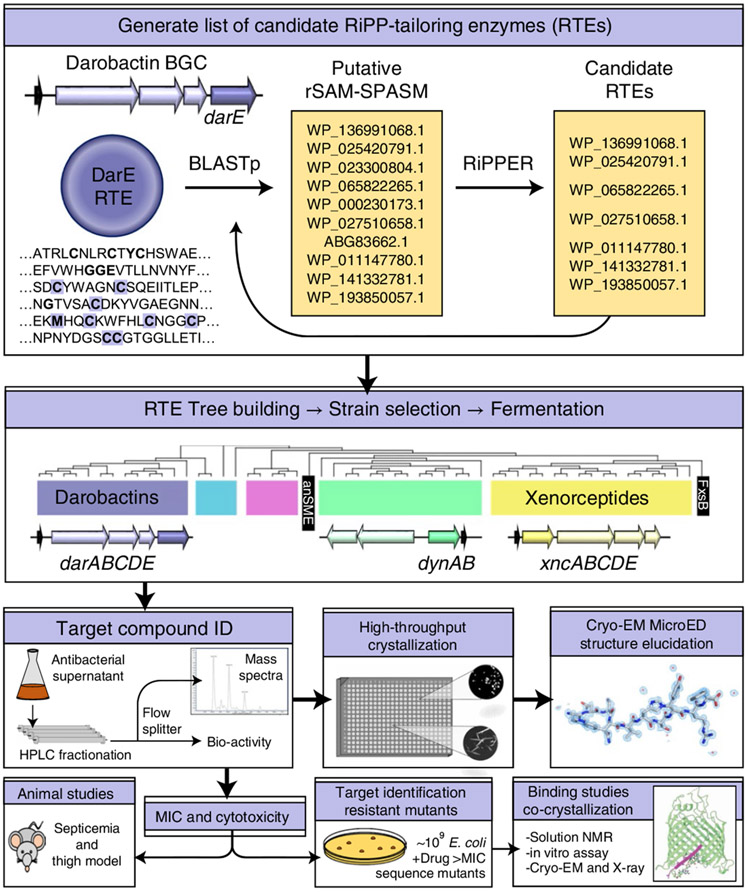
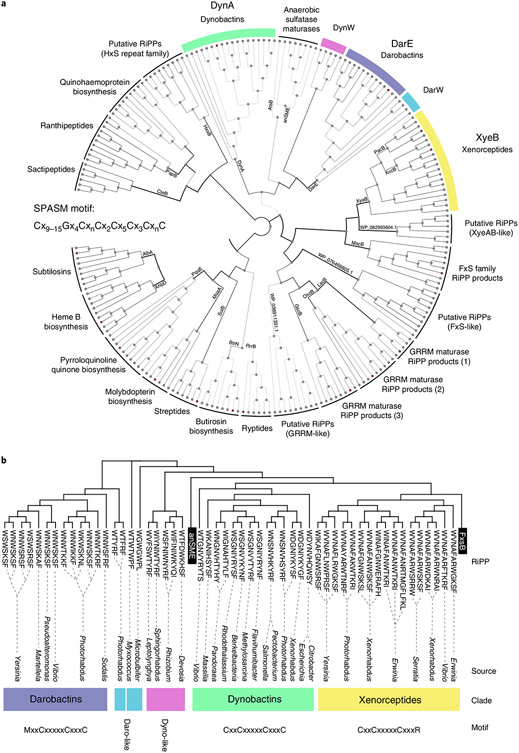
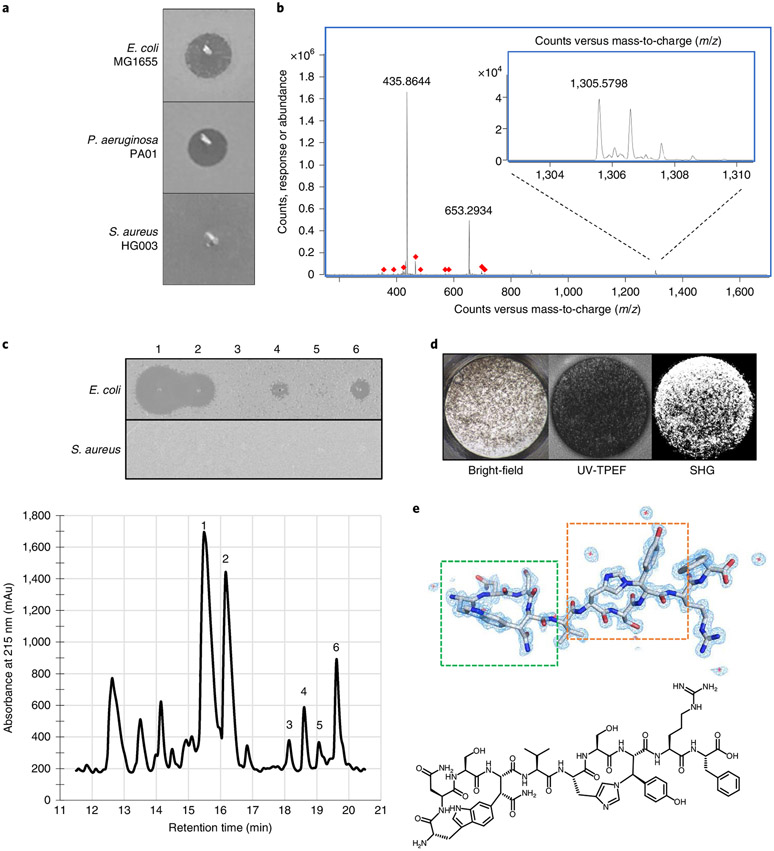
Comment in
-
Computational discovery of dynobactin antibiotics.Nat Microbiol. 2022 Oct;7(10):1512-1513. doi: 10.1038/s41564-022-01234-5. Nat Microbiol. 2022. PMID: 36163502 No abstract available.
References
-
- Brown ED & Wright GD Antibacterial drug discovery in the resistance era. Nature 529, 336–343 (2016). - PubMed
-
- Lewis K The science of antibiotic discovery. Cell 181, 29–45 (2020). - PubMed
-
- Tacconelli E et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis 18, 318–327 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
