

A critical role of calcineurin in stress responses, hyphal formation, and virulence of the pathogenic fungus Trichosporon asahii
- PMID: 36167890
- PMCID: PMC9515189
- DOI: 10.1038/s41598-022-20507-x
A critical role of calcineurin in stress responses, hyphal formation, and virulence of the pathogenic fungus Trichosporon asahii
Abstract
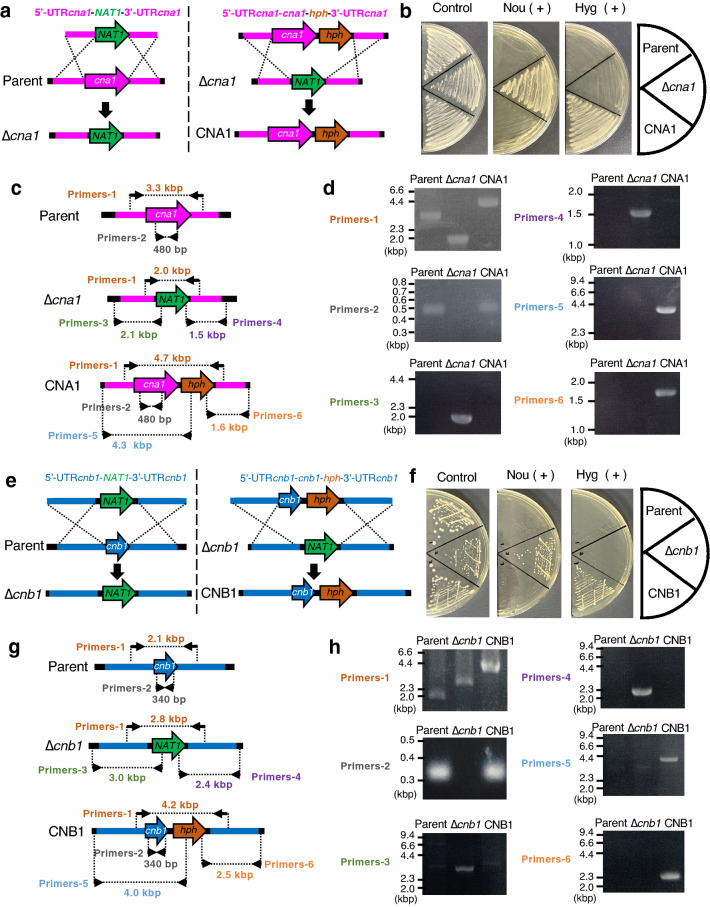
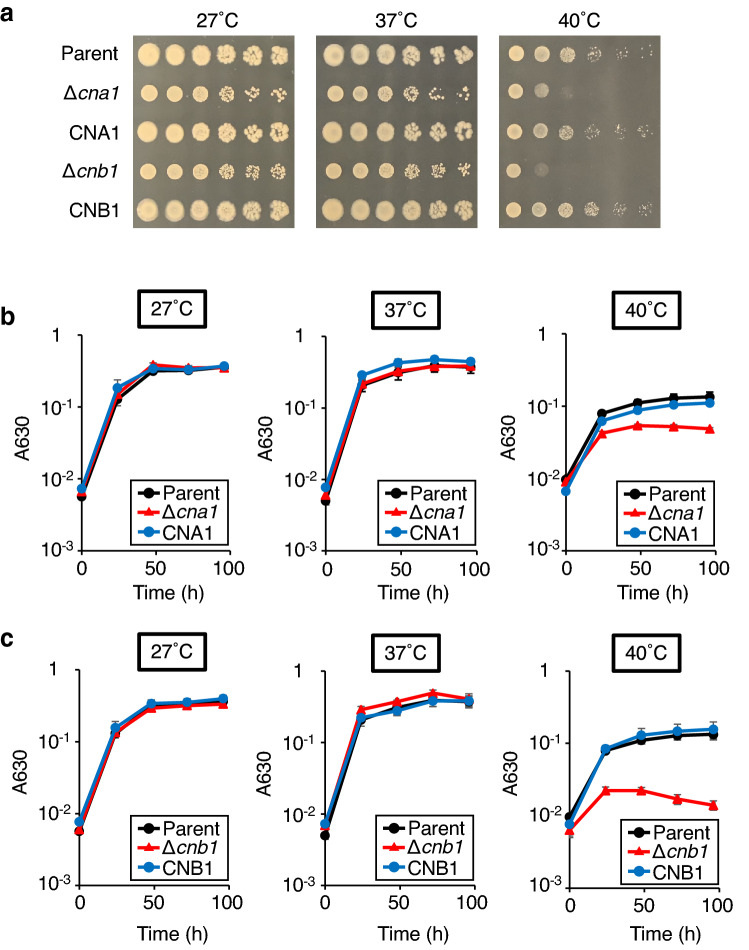
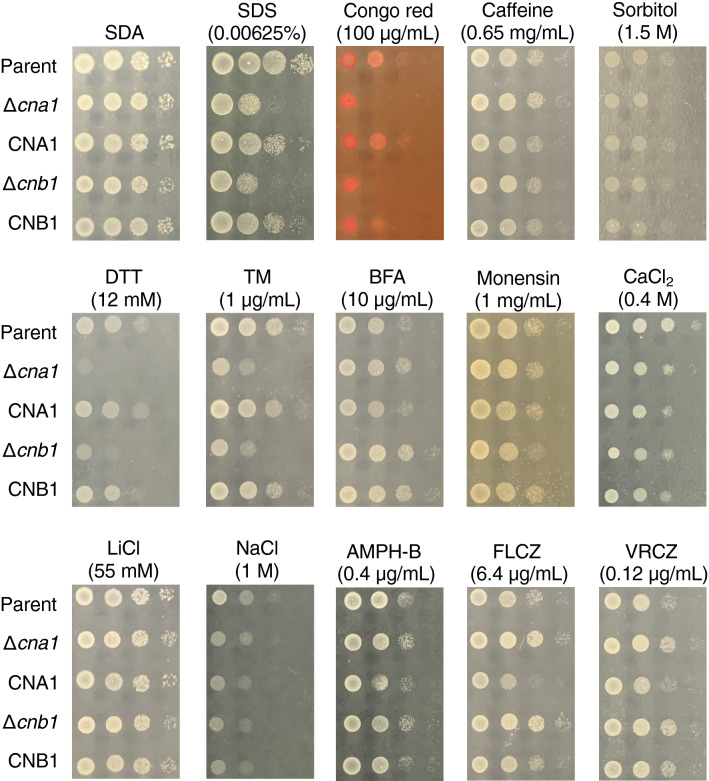
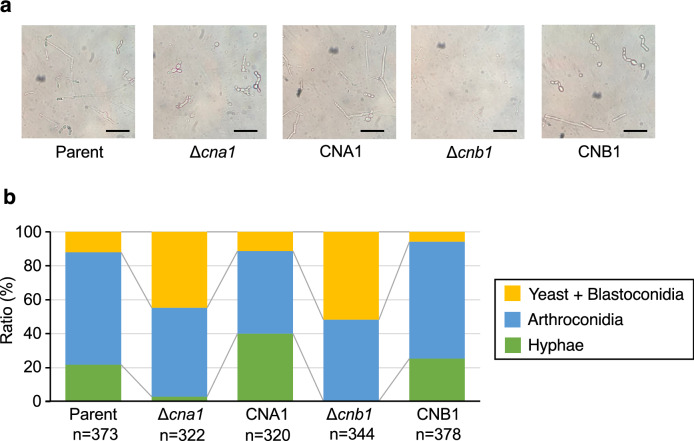
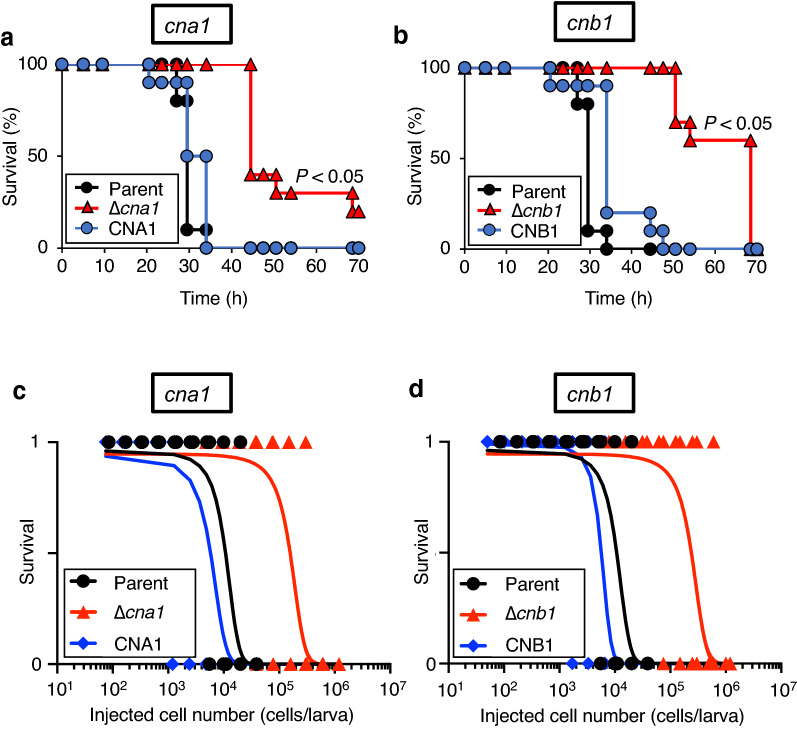
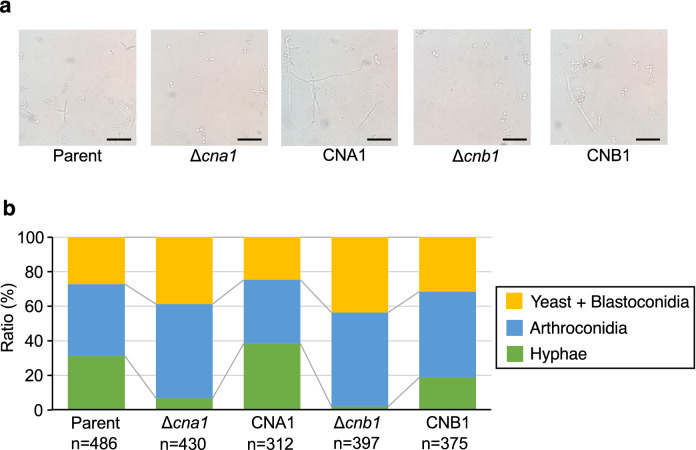
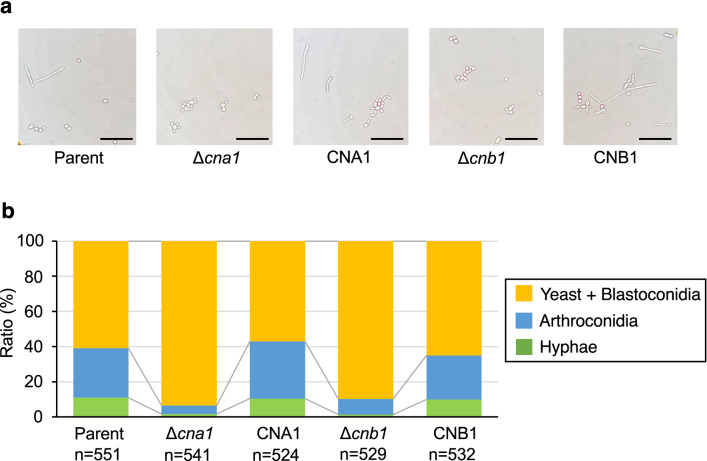
Trichosporon asahii is a conditional pathogenic fungus that causes severe and sometimes fatal infections in immunocompromised patients. While calcineurin, an essential component of a calcium-dependent signaling pathway, is known to regulate stress resistance and virulence of some pathogenic fungi, its role in T. asahii has not been investigated. Here, we demonstrated that calcineurin gene-deficient T. asahii mutants are sensitive to high temperature as well as cell-membrane and cell-wall stress, and exhibit decreased hyphal formation and virulence against silkworms. Growth of T. asahii mutants deficient in genes encoding subunits of calcineurin, cna1 and cnb1, was delayed at 40 °C. The cna1 and cnb1 gene-deficient mutants also showed sensitivity to sodium dodecyl sulfate, Congo red, dithiothreitol, and tunicamycin. On the other hand, these mutants exhibited no sensitivity to caffeine, sorbitol, monensin, CaCl2, LiCl, NaCl, amphotericin B, fluconazole, or voriconazole. The ratio of hyphal formation in the cna1 and cnb1 gene-deficient mutants was decreased. Moreover, the virulence of the cna1 and cnb1 gene-deficient mutants against silkworms was attenuated. These phenotypes were restored by re-introducing each respective gene into the gene-deficient mutants. Our findings suggest that calcineurin has a role in regulating the cellular stress response and virulence of T. asahii.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Hog1-mediated stress tolerance in the pathogenic fungus Trichosporon asahii.Sci Rep. 2023 Aug 19;13(1):13539. doi: 10.1038/s41598-023-40825-y. Sci Rep. 2023. PMID: 37598230 Free PMC article.
-
Tacrolimus inhibits stress responses and hyphal formation via the calcineurin signaling pathway in Trichosporon asahii.Microbiol Immunol. 2023 Feb;67(2):49-57. doi: 10.1111/1348-0421.13039. Epub 2022 Dec 3. Microbiol Immunol. 2023. PMID: 36398783
-
A novel silkworm infection model with fluorescence imaging using transgenic Trichosporon asahii expressing eGFP.Sci Rep. 2020 Jul 3;10(1):10991. doi: 10.1038/s41598-020-67841-6. Sci Rep. 2020. PMID: 32620930 Free PMC article.
-
Strategy to Identify Virulence-Related Genes of the Pathogenic Fungus Trichosporon asahii Using an Efficient Gene-Targeting System.Microbiol Immunol. 2025 Feb;69(2):77-84. doi: 10.1111/1348-0421.13192. Epub 2024 Dec 11. Microbiol Immunol. 2025. PMID: 39660720 Review.
-
Epidemiological study of Trichosporon asahii infections over the past 23 years.Epidemiol Infect. 2020 Jul 24;148:e169. doi: 10.1017/S0950268820001624. Epidemiol Infect. 2020. PMID: 32703332 Free PMC article.
Cited by
-
Adhesion and biofilm formation by two clinical isolates of Trichosporon Cutaneum in various environmental conditions.Braz J Microbiol. 2024 Jun;55(2):1793-1800. doi: 10.1007/s42770-024-01321-1. Epub 2024 Apr 16. Braz J Microbiol. 2024. PMID: 38625517 Free PMC article.
-
Characterization of Virulence Factors, Cellular Stress Response, and Antifungal Susceptibility Testing of Trichosporon spp. Isolated from Northeast Brazilian Patients.J Fungi (Basel). 2025 Mar 26;11(4):255. doi: 10.3390/jof11040255. J Fungi (Basel). 2025. PMID: 40278076 Free PMC article.
-
Hog1-mediated stress tolerance in the pathogenic fungus Trichosporon asahii.Sci Rep. 2023 Aug 19;13(1):13539. doi: 10.1038/s41598-023-40825-y. Sci Rep. 2023. PMID: 37598230 Free PMC article.
-
Metabolome and Transcriptome Combinatory Profiling Reveals Fluconazole Resistance Mechanisms of Trichosporon asahii and the Role of Farnesol in Fluconazole Tolerance.Microorganisms. 2023 Nov 17;11(11):2798. doi: 10.3390/microorganisms11112798. Microorganisms. 2023. PMID: 38004810 Free PMC article.
-
Trichosporon asahii: emerging challenges in pathogenesis and drug resistance.Future Microbiol. 2025 Mar;20(4):333-343. doi: 10.1080/17460913.2025.2457858. Epub 2025 Jan 27. Future Microbiol. 2025. PMID: 39871602 Review.
References
-
- Sugita T, et al. Genetic diversity and biochemical characteristics of Trichosporon asahii isolated from clinical specimens, houses of patients with summer-type-hypersensitivity pneumonitis, and environmental materials. J. Clin. Microbiol. 2001;39:2405–2411. doi: 10.1128/JCM.39.7.2405-2411.2001. - DOI - PMC - PubMed
-
- Zhang E, Sugita T, Tsuboi R, Yamazaki T, Makimura K. The opportunistic yeast pathogen Trichosporon asahii colonizes the skin of healthy individuals: Analysis of 380 healthy individuals by age and gender using a nested polymerase chain reaction assay. Microbiol. Immunol. 2011;55:483–488. doi: 10.1111/j.1348-0421.2011.00341.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
