

Distinct mechanisms of planar polarization by the core and Fat-Dachsous planar polarity pathways in the Drosophila wing
- PMID: 36170824
- PMCID: PMC9631118
- DOI: 10.1016/j.celrep.2022.111419
Distinct mechanisms of planar polarization by the core and Fat-Dachsous planar polarity pathways in the Drosophila wing
Abstract
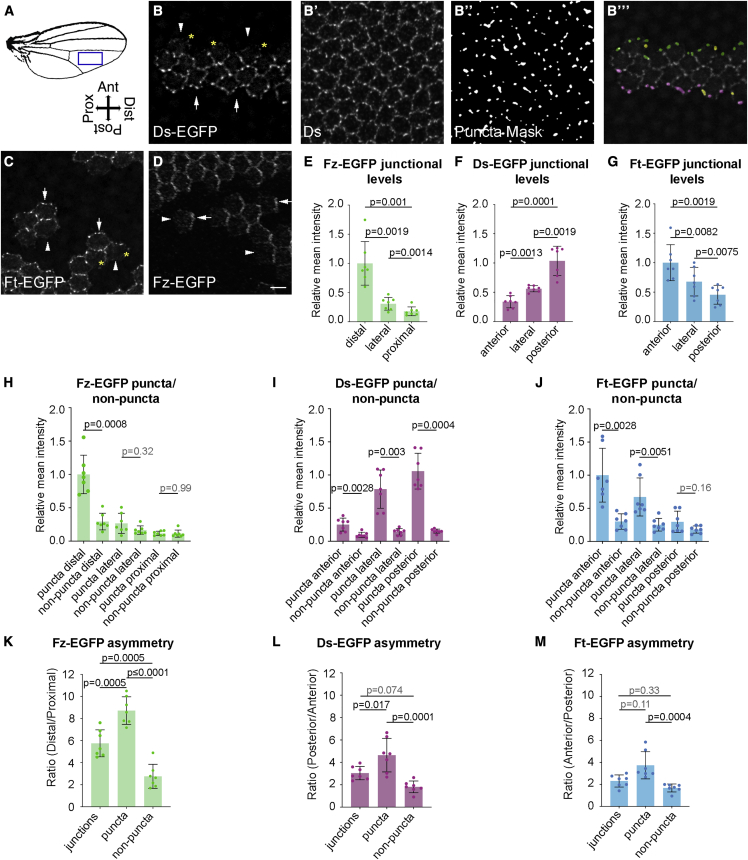
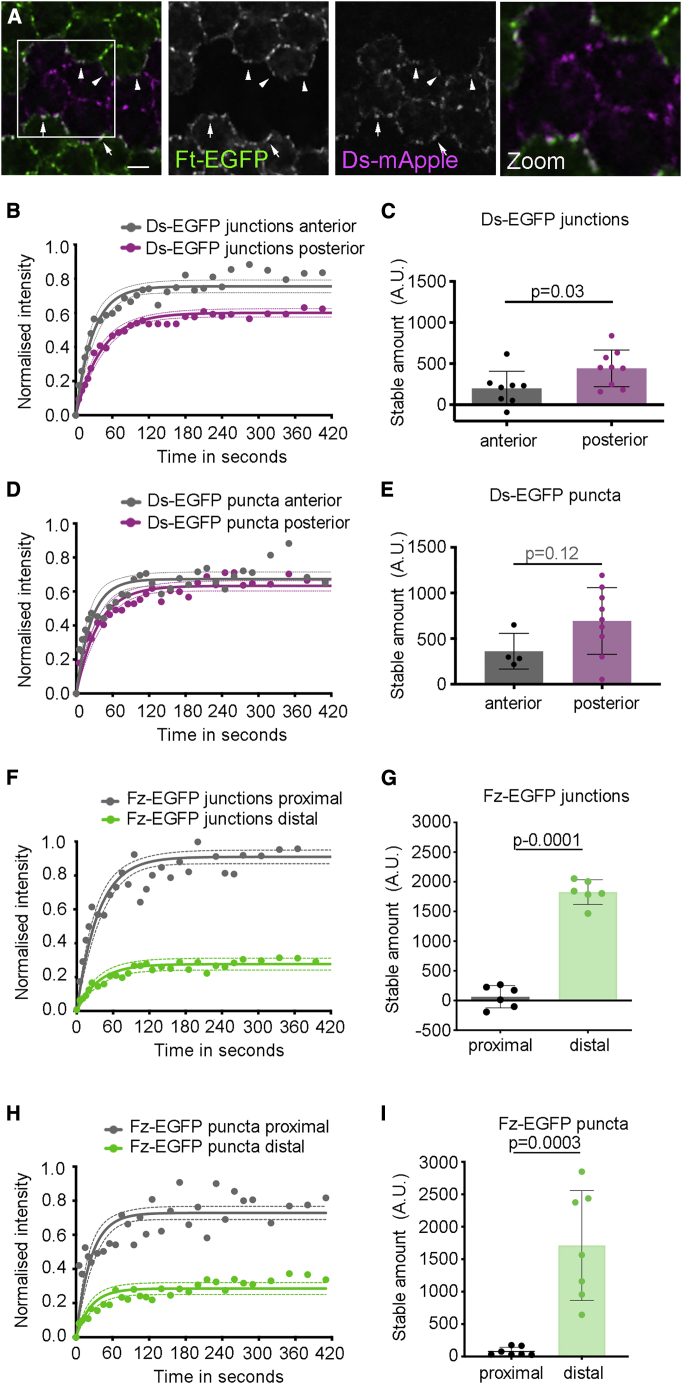
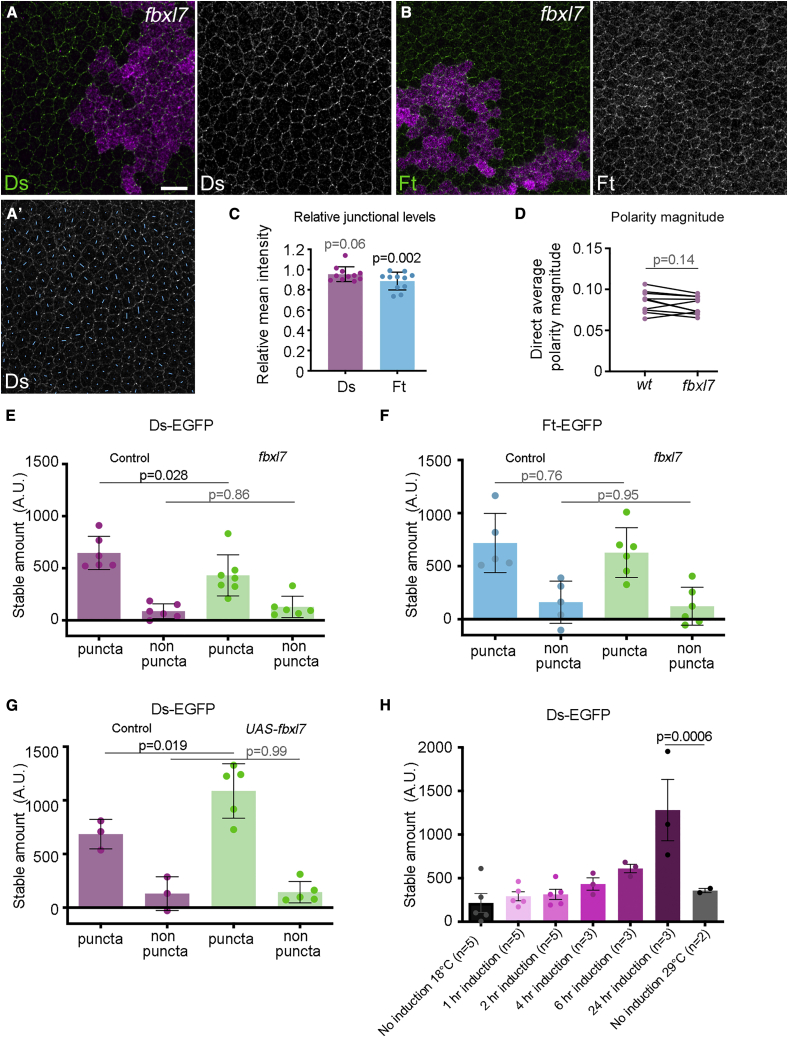
Planar polarity describes the coordinated polarization of cells within a tissue plane, and in animals can be determined by the "core" or Fat-Dachsous pathways. Current models for planar polarity establishment involve two components: tissue-level "global" cues that determine the overall axis of polarity and cell-level feedback-mediated cellular polarity amplification. Here, we investigate the contributions of global cues versus cellular feedback amplification in the core and Fat-Dachsous pathways during Drosophila pupal wing development. We present evidence that these pathways generate planar polarity via distinct mechanisms. Core pathway function is consistent with strong feedback capable of self-organizing cell polarity, which can then be aligned with the tissue axis via weak or transient global cues. Conversely, generation of cell polarity by the Ft-Ds pathway depends on strong global cues in the form of graded patterns of gene expression, which can then be amplified by weak feedback mechanisms.
Keywords: CP: Developmental biology; Dachsous; Fat; Frizzled; PCP; planar cell polarity; planar polarity.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
