

Ventral hippocampus-lateral septum circuitry promotes foraging-related memory
- PMID: 36170832
- PMCID: PMC9605732
- DOI: 10.1016/j.celrep.2022.111402
Ventral hippocampus-lateral septum circuitry promotes foraging-related memory
Abstract
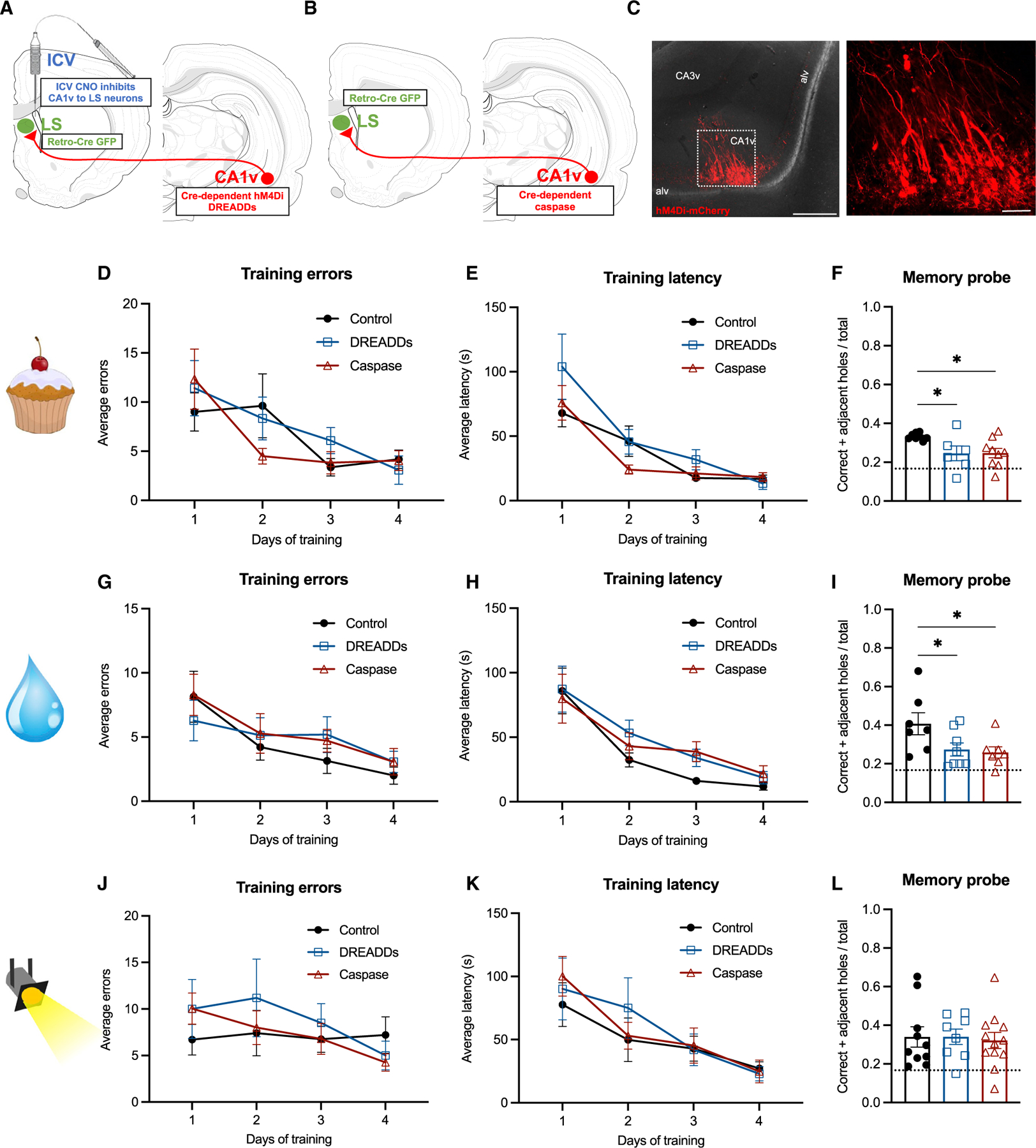
Remembering the location of a food or water source is essential for survival. Here, we reveal that spatial memory for food location is reflected in ventral hippocampus (HPCv) neuron activity and is impaired by HPCv lesion. HPCv mediation of foraging-related memory involves communication to the lateral septum (LS), as either reversible or chronic disconnection of HPCv-to-LS signaling impairs spatial memory retention for food or water location. This neural pathway selectively encodes appetitive spatial memory, as HPCv-LS disconnection does not affect spatial memory for escape location in a negative reinforcement procedure, food intake, or social and olfactory-based appetitive learning. Neural pathway tracing and functional mapping analyses reveal that LS neurons recruited during the appetitive spatial memory procedure are primarily GABAergic neurons that project to the lateral hypothalamus. Collective results emphasize that the neural substrates controlling spatial memory are outcome specific based on reinforcer modality.
Keywords: CP: Neuroscience; feeding; foraging; lateral hypothalamus; motivation; prefrontal cortex; reward.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




Similar articles
-
Ventral hippocampus neurons encode meal-related memory.Nat Commun. 2025 Jun 11;16(1):4898. doi: 10.1038/s41467-025-59687-1. Nat Commun. 2025. PMID: 40500290 Free PMC article.
-
Ventral hippocampus neurons encode meal-related memory.bioRxiv [Preprint]. 2023 Oct 10:2023.10.10.561731. doi: 10.1101/2023.10.10.561731. bioRxiv. 2023. Update in: Nat Commun. 2025 Jun 11;16(1):4898. doi: 10.1038/s41467-025-59687-1. PMID: 37873229 Free PMC article. Updated. Preprint.
-
Conjunctive spatial and self-motion codes are topographically organized in the GABAergic cells of the lateral septum.PLoS Biol. 2021 Aug 30;19(8):e3001383. doi: 10.1371/journal.pbio.3001383. eCollection 2021 Aug. PLoS Biol. 2021. PMID: 34460812 Free PMC article.
-
Putting Together Pieces of the Lateral Septum: Multifaceted Functions and Its Neural Pathways.eNeuro. 2021 Dec 3;8(6):ENEURO.0315-21.2021. doi: 10.1523/ENEURO.0315-21.2021. Print 2021 Nov-Dec. eNeuro. 2021. PMID: 34764187 Free PMC article. Review.
-
Lateral septum as a nexus for mood, motivation, and movement.Neurosci Biobehav Rev. 2021 Jul;126:544-559. doi: 10.1016/j.neubiorev.2021.03.029. Epub 2021 Apr 10. Neurosci Biobehav Rev. 2021. PMID: 33848512 Review.
Cited by
-
An ultra-short-acting benzodiazepine in thalamic nucleus reuniens undermines fear extinction via intermediation of hippocamposeptal circuits.Commun Biol. 2024 Jun 14;7(1):728. doi: 10.1038/s42003-024-06417-w. Commun Biol. 2024. PMID: 38877285 Free PMC article.
-
A thalamic nucleus reuniens-lateral septum-lateral hypothalamus circuit for comorbid anxiety-like behaviors in chronic itch.Sci Adv. 2024 Aug 16;10(33):eadn6272. doi: 10.1126/sciadv.adn6272. Epub 2024 Aug 16. Sci Adv. 2024. PMID: 39150998 Free PMC article.
-
Western diet consumption impairs memory function via dysregulated hippocampus acetylcholine signaling.Brain Behav Immun. 2024 May;118:408-422. doi: 10.1016/j.bbi.2024.03.015. Epub 2024 Mar 8. Brain Behav Immun. 2024. PMID: 38461956 Free PMC article.
-
Evidence for novelty reward cross-cueing in the odor span task in rats: implications for odor-based reward-motivated tasks.Learn Mem. 2024 Jan 29;31(1-2):a053871. doi: 10.1101/lm.053871.123. Print 2024 Jan-Feb. Learn Mem. 2024. PMID: 38286523 Free PMC article.
-
Ventral hippocampus neurons encode meal-related memory.Nat Commun. 2025 Jun 11;16(1):4898. doi: 10.1038/s41467-025-59687-1. Nat Commun. 2025. PMID: 40500290 Free PMC article.
References
-
- Barnes CA (1979). Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J. Comp. Physiol. Psychol 93, 74–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
