

Integrated single-cell RNA-seq analysis identifies immune heterogeneity associated with KRAS/TP53 mutation status and tumor-sideness in colorectal cancers
- PMID: 36172359
- PMCID: PMC9510840
- DOI: 10.3389/fimmu.2022.961350
Integrated single-cell RNA-seq analysis identifies immune heterogeneity associated with KRAS/TP53 mutation status and tumor-sideness in colorectal cancers
Abstract
Background: The main objective of this study was to analyze the effects of KRAS/TP53 mutation status and tumor sideness on the immune microenvironment of colorectal cancer using integrated scRNA-seq data.
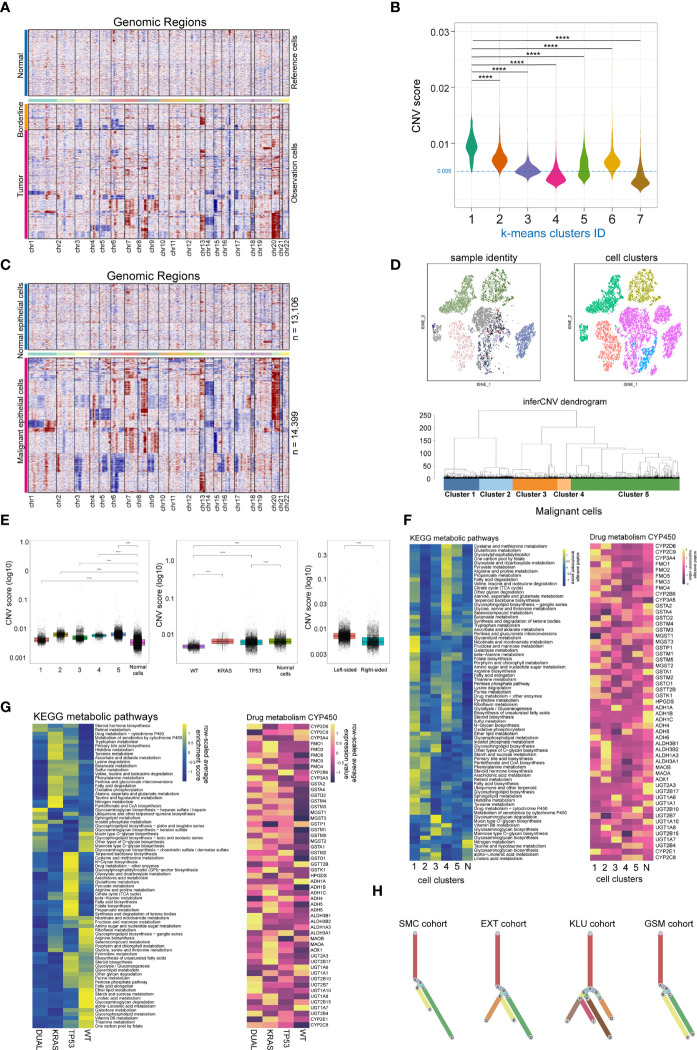
Methods: A total of 78 scRNA-seq datasets, comprising 42 treatment-naive colorectal tumors, 13 tumor adjacent tissues and 23 normal mucosa tissues were included. Standardized Seurat procedures were applied to identify cellular components with canonical cell marks. The batch-effect was assessed and corrected using harmony algorithm. The scMetabolism algorithm was used for single-cell metabolic analysis. The results and clinical significance were further validated using immunofluorescent-staining and TCGA-COAD datasets. Immune-infiltration scores of bulk-RNA-seq data were estimated using ssGSEA. The presto-wilcoxauc algorithm was used to identify differentially enriched genes or pathways across different subgroups. Two-sided p-value less than 0.05 was considered statistically significant.
Results: We refined the landscape of functional immune cell subtypes, especially T cells and myeloid cells, across normal mucosa, tumor adjacent and tumor tissue. The existence and function of two states of exhausted CD8+ T (Tex) subtypes in colorectal cancer, and FOLR2+ LYVE1+ macrophages indicating unfavorable prognosis in colorectal cancer were identified and validated. The diverse tumor mutation status reshaped the immune cell function and immune checkpoint ligands/receptors (ICLs/ICRs) expression pattern. Importantly, the KRAS/TP53 dual mutations significantly reduced the major energy metabolic functions in immune cells, and promoted the cell-to-cell communications towards immunosuppression in colorectal cancers. The results revealed LAG3, CD24-SIGLEC10 and HBEGF-CD9 pathways as potential therapeutic targets for dual mutant colorectal cancers.
Conclusions: We revealed that the immune microenvironment underwent a gradual remodeling with an enrichment of immunosuppressive myeloid cells from normal mucosa to tumor regions in colorectal cancers. Moreover, we revealed the metabolic heterogeneity of tumor-infiltrating immune cells and suggested that the KRAS/TP53 dual mutation may impair antitumor immunity by reducing T and myeloid cell energy metabolism and reshaping cellular interactions toward immunosuppression.
Keywords: KRAS mutation; TP53 mutation; clinical prognosis; colorectal cancer; therapeutic targets; tumor immune microenvironment; tumor sideness.
Copyright © 2022 Liu, Xu, Wu, Shan, Wang, Wu, Ding, Huang and Wang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






Similar articles
-
Mutation-Driven Immune Microenvironments in Non-Small Cell Lung Cancer: Unrevealing Patterns through Cluster Analysis.Yonsei Med J. 2024 Dec;65(12):683-694. doi: 10.3349/ymj.2024.0062. Yonsei Med J. 2024. PMID: 39609084 Free PMC article.
-
Comparative analyses of the prognosis, tumor immune microenvironment, and drug treatment response between left-sided and right-sided colon cancer by integrating scRNA-seq and bulk RNA-seq data.Aging (Albany NY). 2023 Jul 21;15(14):7098-7123. doi: 10.18632/aging.204894. Epub 2023 Jul 21. Aging (Albany NY). 2023. PMID: 37480572 Free PMC article.
-
Mathematical modeling the order of driver gene mutations in colorectal cancer.PLoS Comput Biol. 2023 Jun 27;19(6):e1011225. doi: 10.1371/journal.pcbi.1011225. eCollection 2023 Jun. PLoS Comput Biol. 2023. PMID: 37368936 Free PMC article.
-
Impact of KRAS mutation on the tumor microenvironment in colorectal cancer.Int J Biol Sci. 2024 Mar 3;20(5):1947-1964. doi: 10.7150/ijbs.88779. eCollection 2024. Int J Biol Sci. 2024. PMID: 38481800 Free PMC article. Review.
-
KRAS and TP53 mutations in inflammatory bowel disease-associated colorectal cancer: a meta-analysis.Oncotarget. 2017 Mar 28;8(13):22175-22186. doi: 10.18632/oncotarget.14549. Oncotarget. 2017. PMID: 28077799 Free PMC article. Review.
Cited by
-
Single-cell transcriptome sequencing reveals spatial distribution of IL34+ cancer-associated fibroblasts in hepatocellular carcinoma tumor microenvironment.NPJ Precis Oncol. 2023 Dec 11;7(1):133. doi: 10.1038/s41698-023-00483-9. NPJ Precis Oncol. 2023. PMID: 38081923 Free PMC article.
-
Novel tumor-associated macrophage populations and subpopulations by single cell RNA sequencing.Front Immunol. 2024 Jan 29;14:1264774. doi: 10.3389/fimmu.2023.1264774. eCollection 2023. Front Immunol. 2024. PMID: 38347955 Free PMC article. Review.
-
Unveiling the key mechanisms of FOLR2+ macrophage-mediated antitumor immunity in breast cancer using integrated single-cell RNA sequencing and bulk RNA sequencing.Breast Cancer Res. 2025 Mar 5;27(1):31. doi: 10.1186/s13058-025-01980-4. Breast Cancer Res. 2025. PMID: 40045365 Free PMC article.
-
Exploring the Role of Circulating Cell-Free RNA in the Development of Colorectal Cancer.Int J Mol Sci. 2023 Jul 3;24(13):11026. doi: 10.3390/ijms241311026. Int J Mol Sci. 2023. PMID: 37446204 Free PMC article. Review.
-
Unlocking the potential of T-cell metabolism reprogramming: Advancing single-cell approaches for precision immunotherapy in tumour immunity.Clin Transl Med. 2024 Mar;14(3):e1620. doi: 10.1002/ctm2.1620. Clin Transl Med. 2024. PMID: 38468489 Free PMC article. Review.
References
-
- Fokas E, Schlenska-Lange A, Polat B, Klautke G, Grabenbauer G, Fietkau R, et al. . Chemoradiotherapy plus induction or consolidation chemotherapy as total neoadjuvant therapy for patients with locally advanced rectal cancer: Long-term results of the CAO/ARO/AIO-12 randomized clinical trial. JAMA Oncol (2022) 8(1):e215445. doi: 10.1001/jamaoncol.2021.5445 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous