

Repression and 3D-restructuring resolves regulatory conflicts in evolutionarily rearranged genomes
- PMID: 36179666
- PMCID: PMC9567273
- DOI: 10.1016/j.cell.2022.09.006
Repression and 3D-restructuring resolves regulatory conflicts in evolutionarily rearranged genomes
Abstract
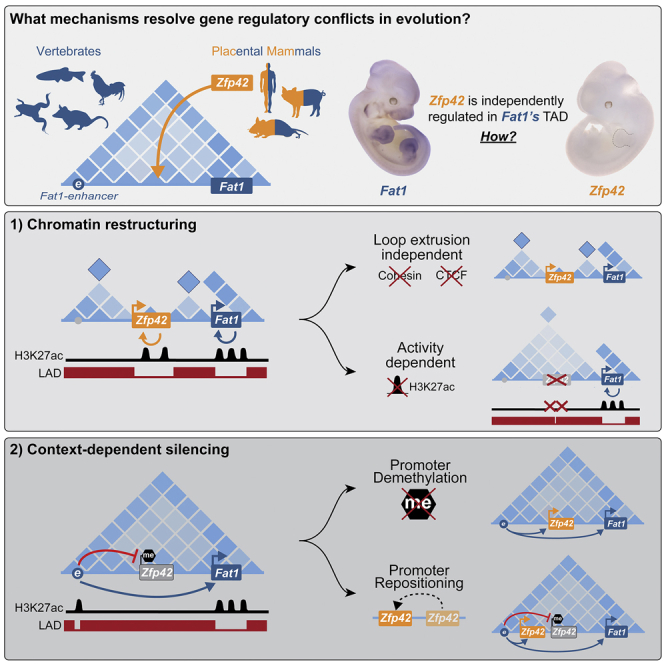
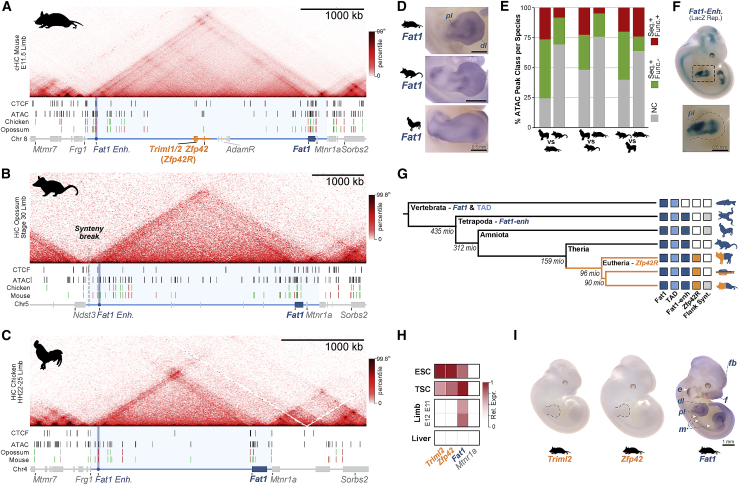
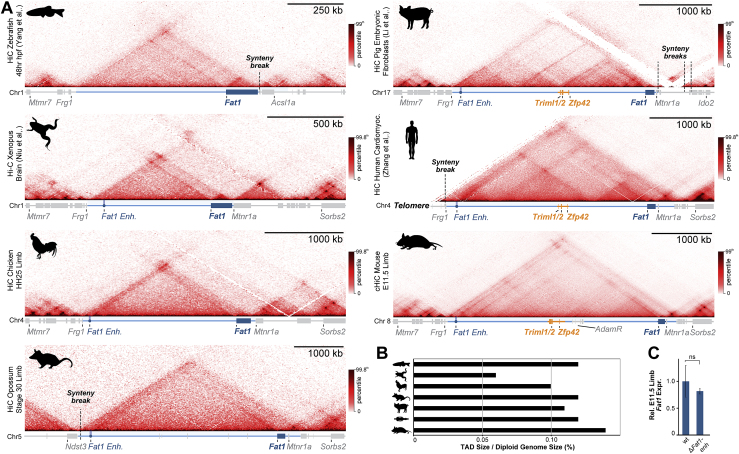
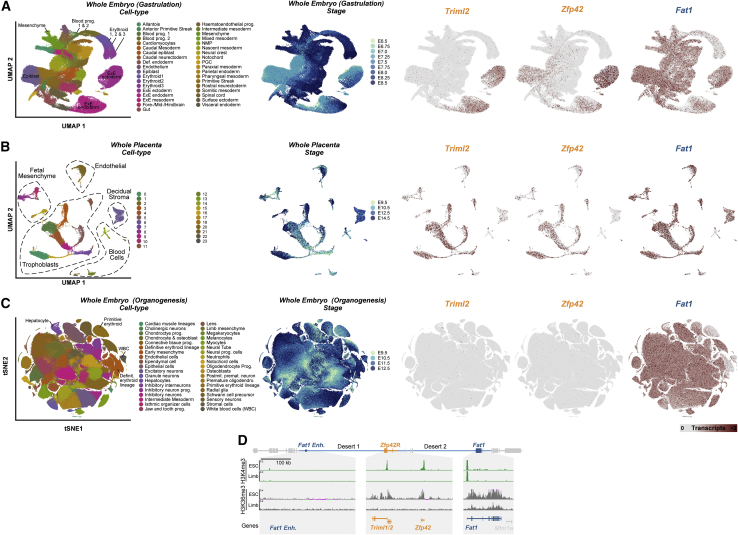
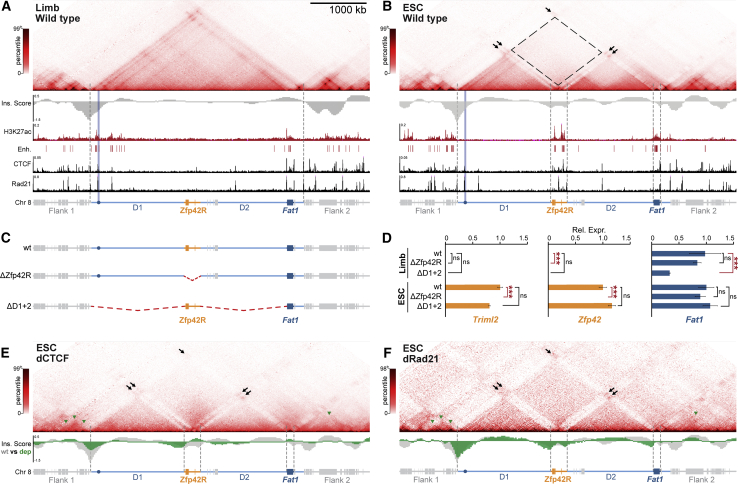
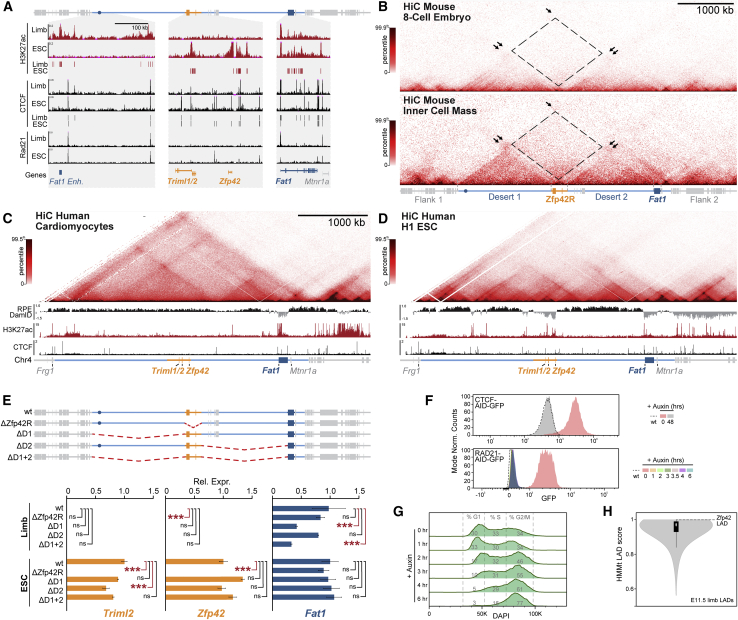
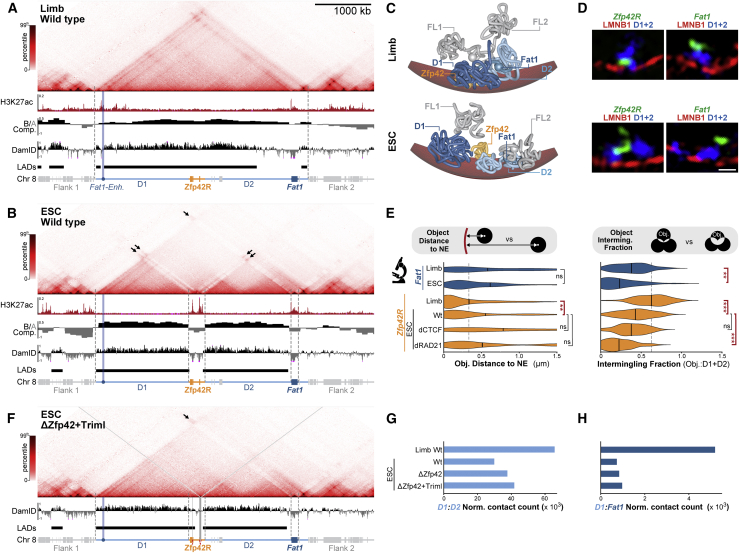
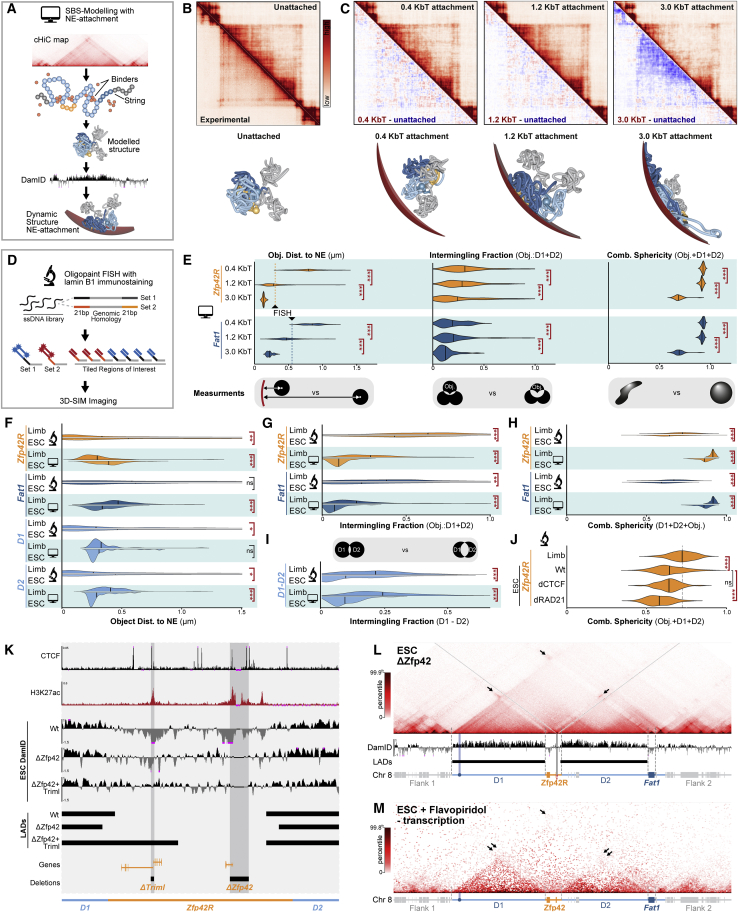
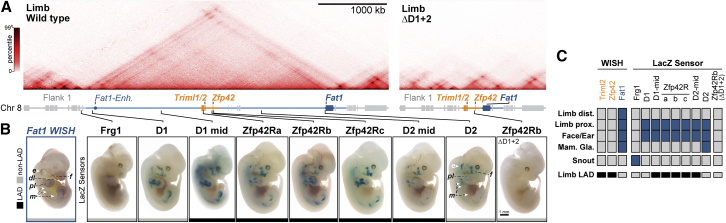
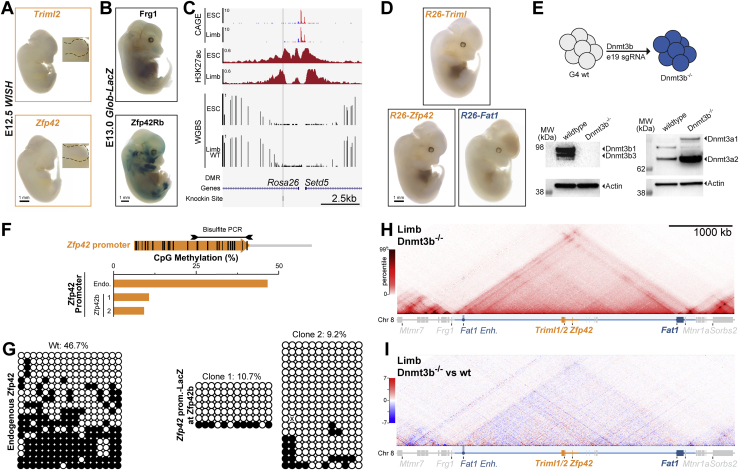
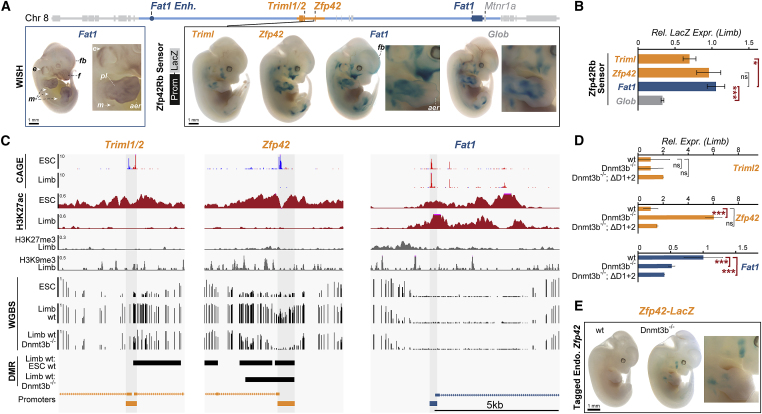
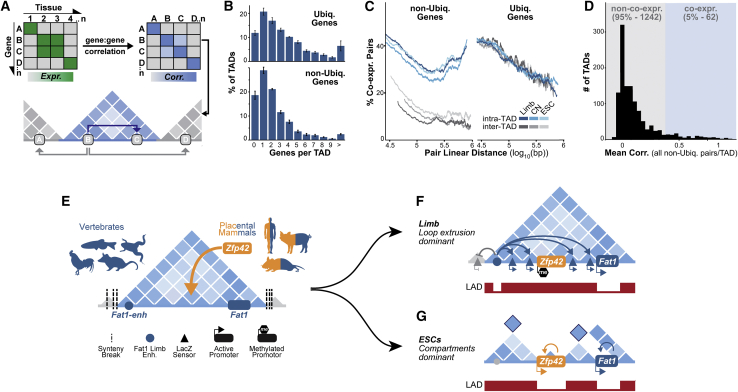
Regulatory landscapes drive complex developmental gene expression, but it remains unclear how their integrity is maintained when incorporating novel genes and functions during evolution. Here, we investigated how a placental mammal-specific gene, Zfp42, emerged in an ancient vertebrate topologically associated domain (TAD) without adopting or disrupting the conserved expression of its gene, Fat1. In ESCs, physical TAD partitioning separates Zfp42 and Fat1 with distinct local enhancers that drive their independent expression. This separation is driven by chromatin activity and not CTCF/cohesin. In contrast, in embryonic limbs, inactive Zfp42 shares Fat1's intact TAD without responding to active Fat1 enhancers. However, neither Fat1 enhancer-incompatibility nor nuclear envelope-attachment account for Zfp42's unresponsiveness. Rather, Zfp42's promoter is rendered inert to enhancers by context-dependent DNA methylation. Thus, diverse mechanisms enabled the integration of independent Zfp42 regulation in the Fat1 locus. Critically, such regulatory complexity appears common in evolution as, genome wide, most TADs contain multiple independently expressed genes.
Keywords: 3D genome organization; CTCF; DNA methylation; cohesin; developmental gene regulation; enhancer-promoter specificity; evolution; lamina-associated domain; loop extrusion; topologically associating domains.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
A TAD refined for gene regulation.Nat Rev Genet. 2022 Dec;23(12):714. doi: 10.1038/s41576-022-00549-7. Nat Rev Genet. 2022. PMID: 36316395 No abstract available.
References
-
- Acemel R.D., Maeso I., Gómez-Skarmeta J.L. Topologically associated domains: a successful scaffold for the evolution of gene regulation in animals. Wiley Interdiscip. Rev. Dev. Biol. 2017;6:e265. - PubMed
-
- Allen M.P., Tildesley D.J. Oxford University Press; 2017. Computer Simulation of Liquids.
-
- Allou L., Balzano S., Magg A., Quinodoz M., Royer-Bertrand B., Schöpflin R., Chan W.L., Speck-Martins C.E., Carvalho D.R., Farage L., et al. Non-coding deletions identify Maenli lncRNA as a limb-specific En1 regulator. Nature. 2021;592:93–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
