

Proper acquisition of cell class identity in organoids allows definition of fate specification programs of the human cerebral cortex
- PMID: 36179669
- PMCID: PMC9990683
- DOI: 10.1016/j.cell.2022.09.010
Proper acquisition of cell class identity in organoids allows definition of fate specification programs of the human cerebral cortex
Abstract
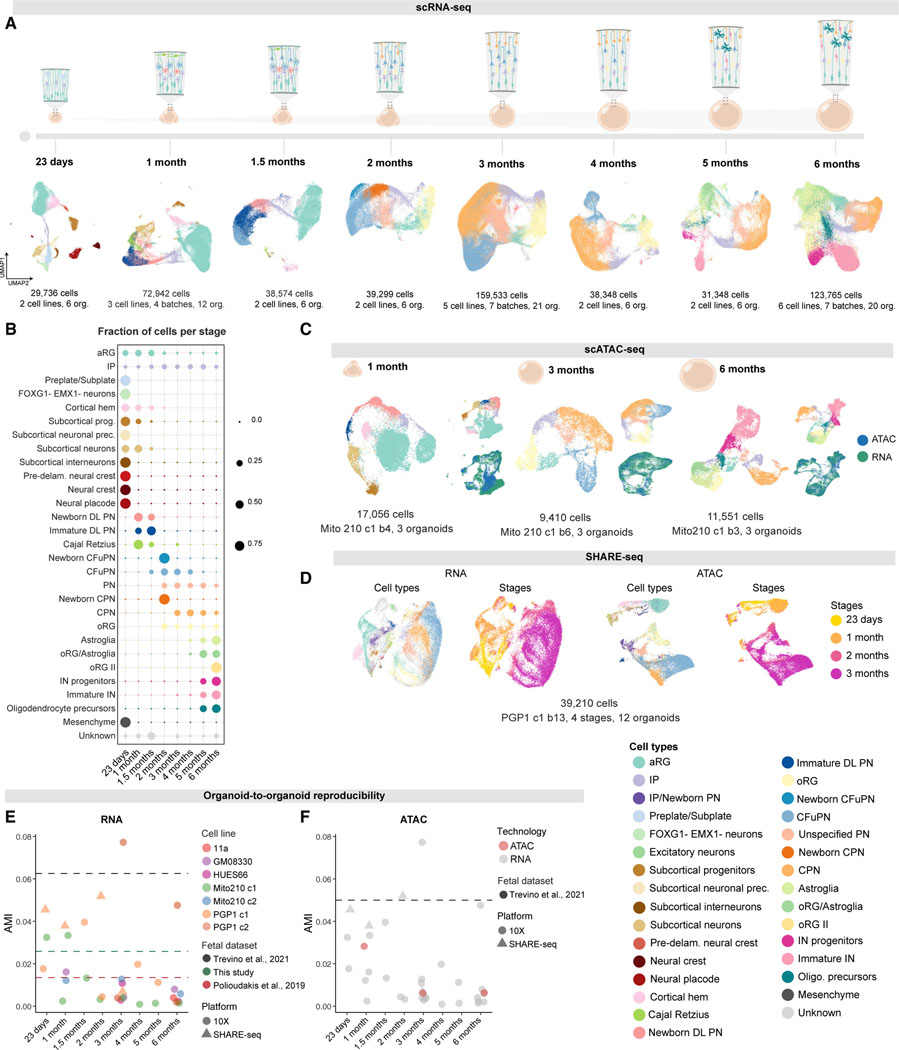
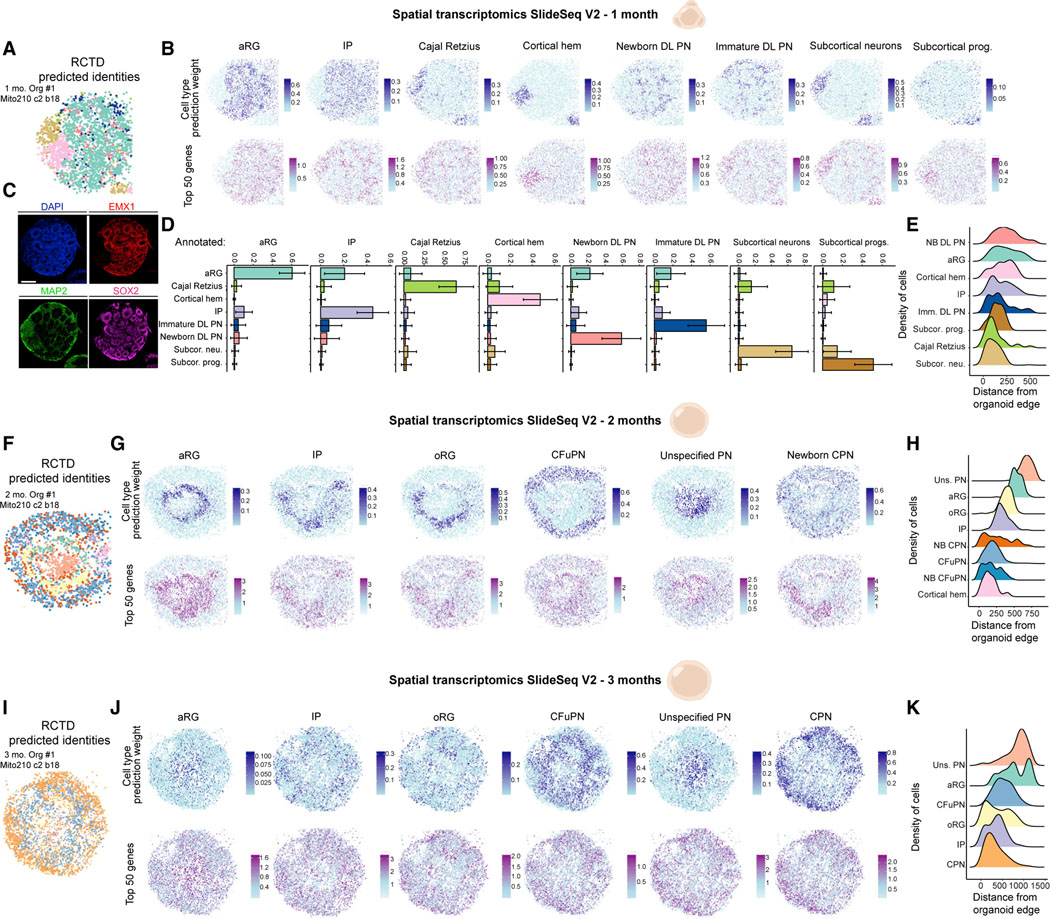
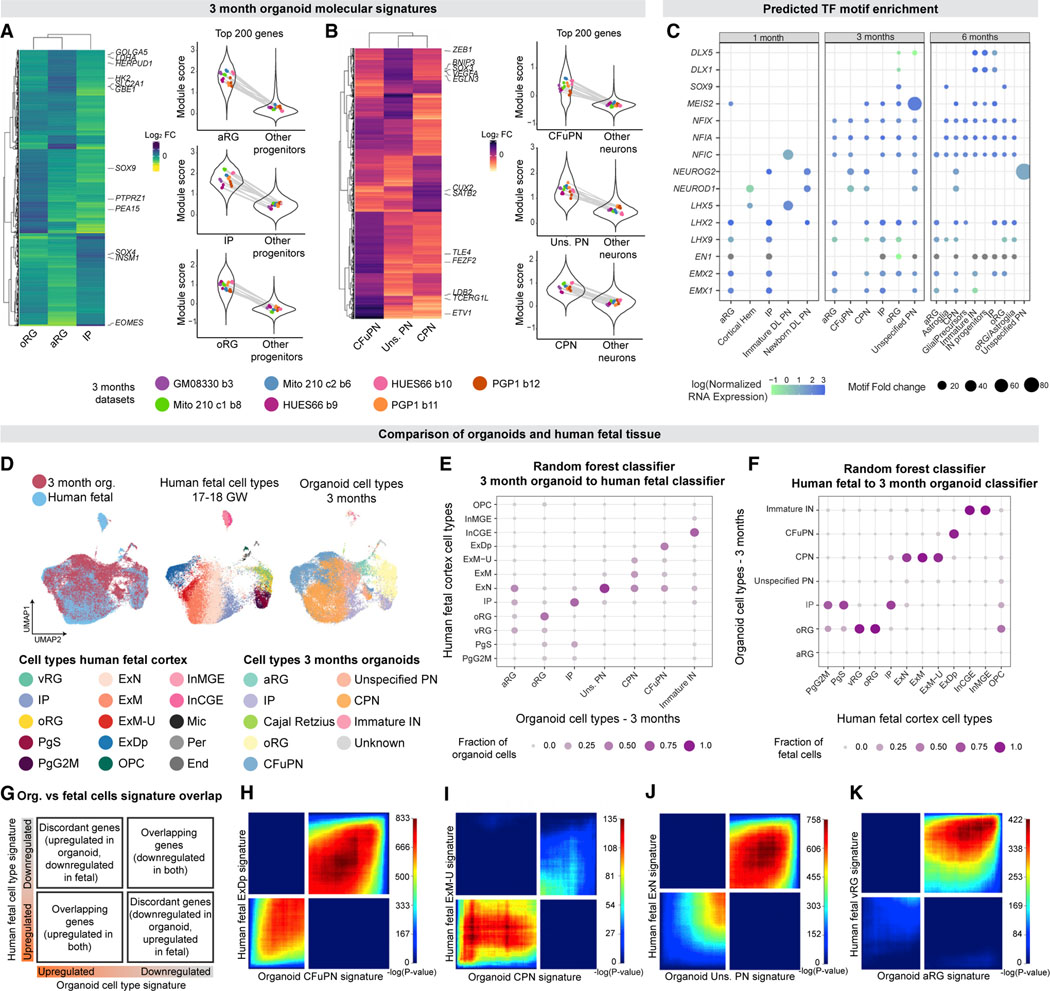
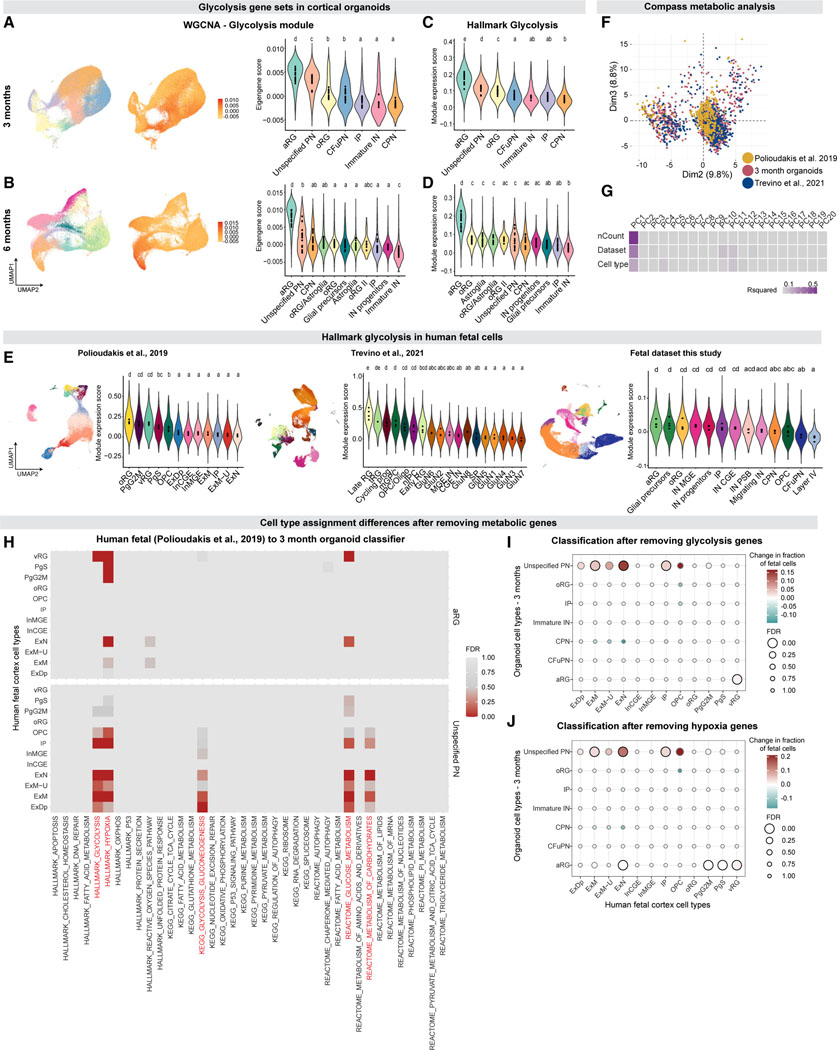
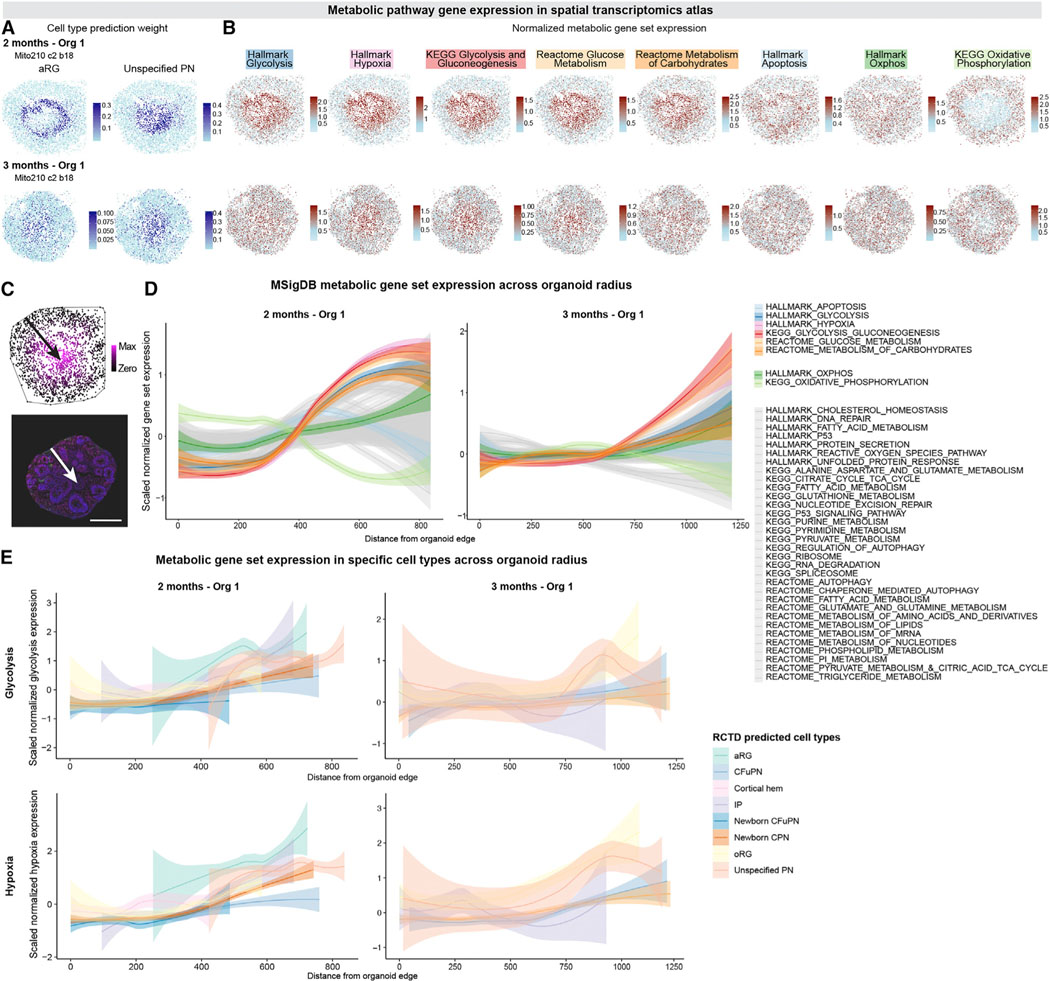
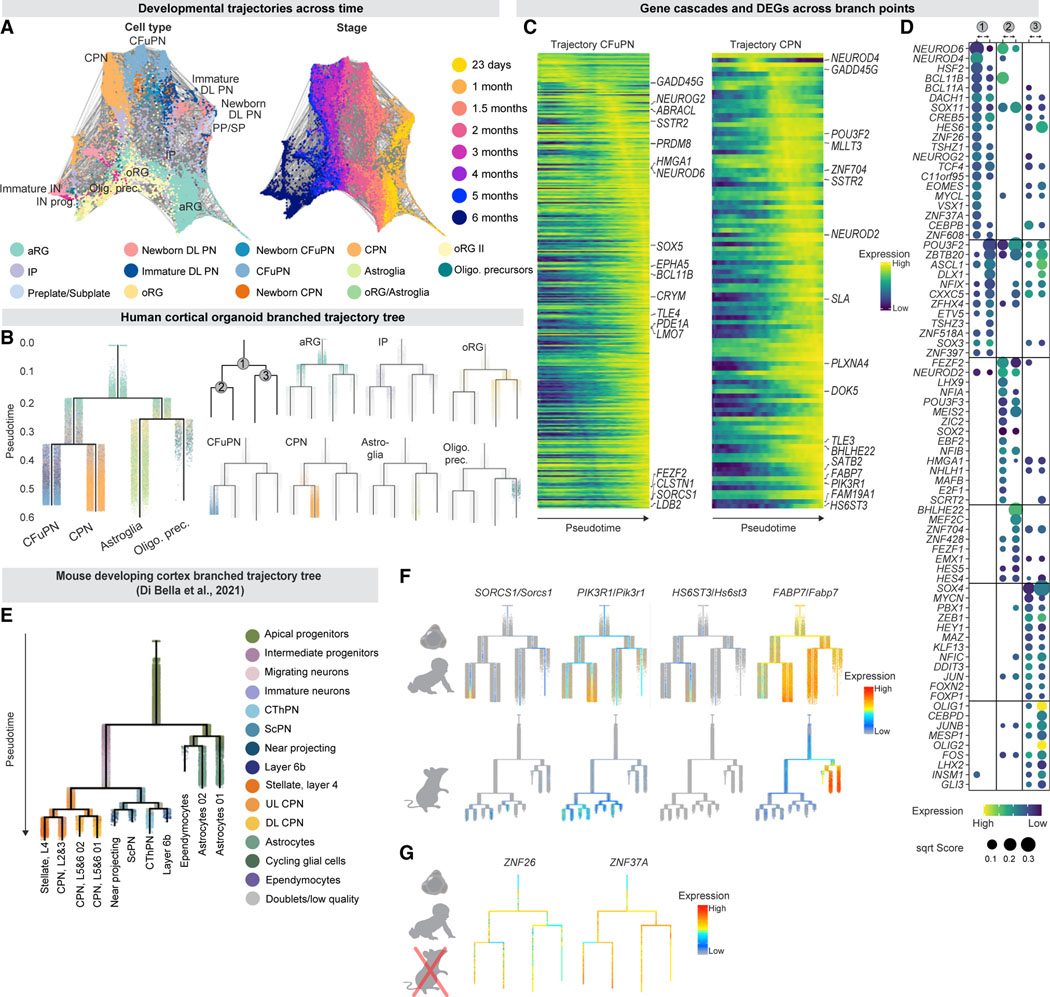
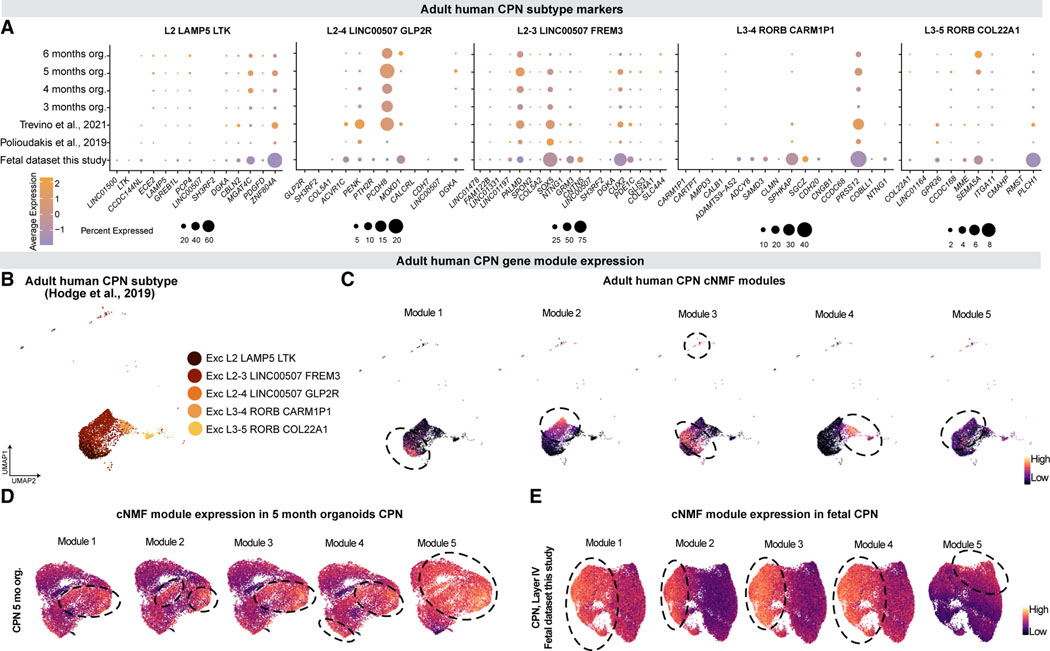
Realizing the full utility of brain organoids to study human development requires understanding whether organoids precisely replicate endogenous cellular and molecular events, particularly since acquisition of cell identity in organoids can be impaired by abnormal metabolic states. We present a comprehensive single-cell transcriptomic, epigenetic, and spatial atlas of human cortical organoid development, comprising over 610,000 cells, from generation of neural progenitors through production of differentiated neuronal and glial subtypes. We show that processes of cellular diversification correlate closely to endogenous ones, irrespective of metabolic state, empowering the use of this atlas to study human fate specification. We define longitudinal molecular trajectories of cortical cell types during organoid development, identify genes with predicted human-specific roles in lineage establishment, and uncover early transcriptional diversity of human callosal neurons. The findings validate this comprehensive atlas of human corticogenesis in vitro as a resource to prime investigation into the mechanisms of human cortical development.
Keywords: Brain organoids; Cortical development; In vitro metabolism; Multiomics; Neuronal diversity; Single cell RNA-seq; Spatial transcriptomics.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests P.A. is a SAB member at Herophilus, Rumi Therapeutics, and Foresite Labs, and is a co-founder of Vesalius and a co-founder and equity holder at Foresite Labs. A.R. is a founder and equity holder of Celsius Therapeutics, an equity holder in Immunitas Therapeutics and until August 31, 2020, was a SAB member of Syros Pharmaceuticals, Neogene Therapeutics, Asimov and Thermo Fisher Scientific. From August 1, 2020, A.R. has been an employee of Genentech. M.P. is an employee of Roche.
Figures
References
-
- Betizeau M, Cortay V, Patti D, Pfister S, Gautier E, Bellemin-Ménard A, Afanassieff M, Huissoud C, Douglas RJ, Kennedy H, and Dehay C. (2013). Precursor diversity and complexity of lineage relationships in the outer subventricular zone of the primate. Neuron 80, 442–457. 10.1016/j.neuron.2013.09.032. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
