Pharmacological targeting of G protein-coupled receptor heteromers
- PMID: 36182040
- PMCID: PMC9645299
- DOI: 10.1016/j.phrs.2022.106476
Pharmacological targeting of G protein-coupled receptor heteromers
Abstract
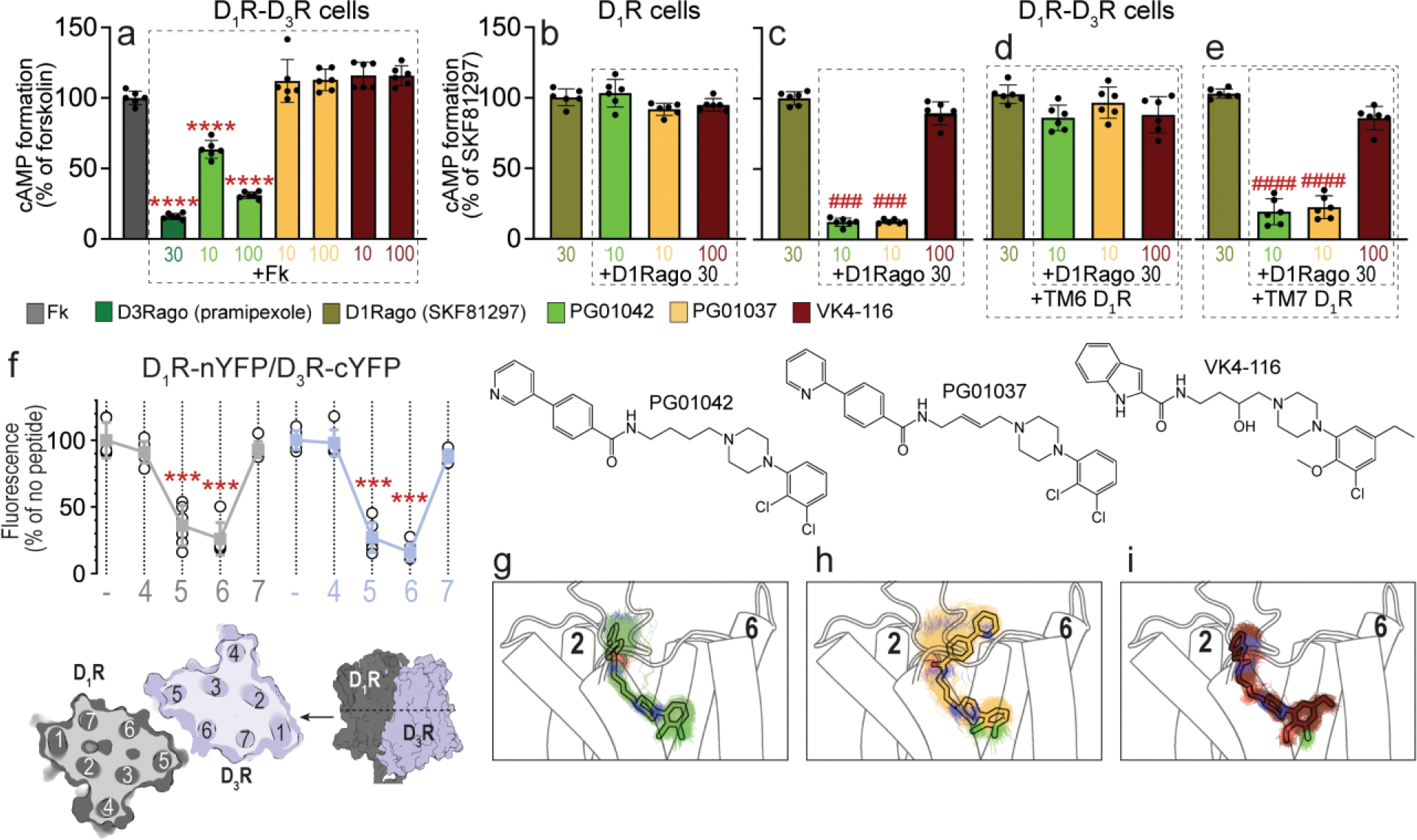
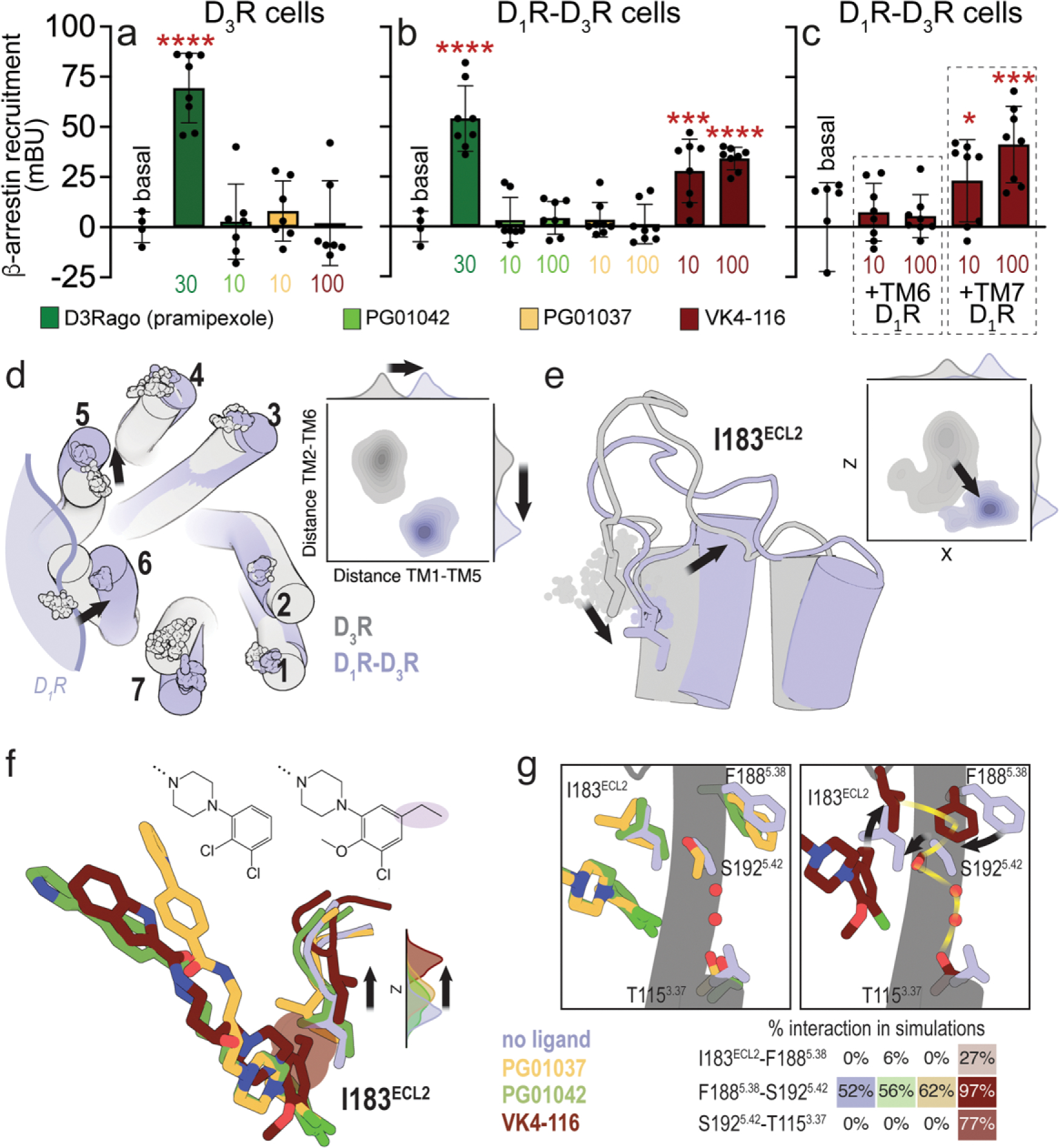
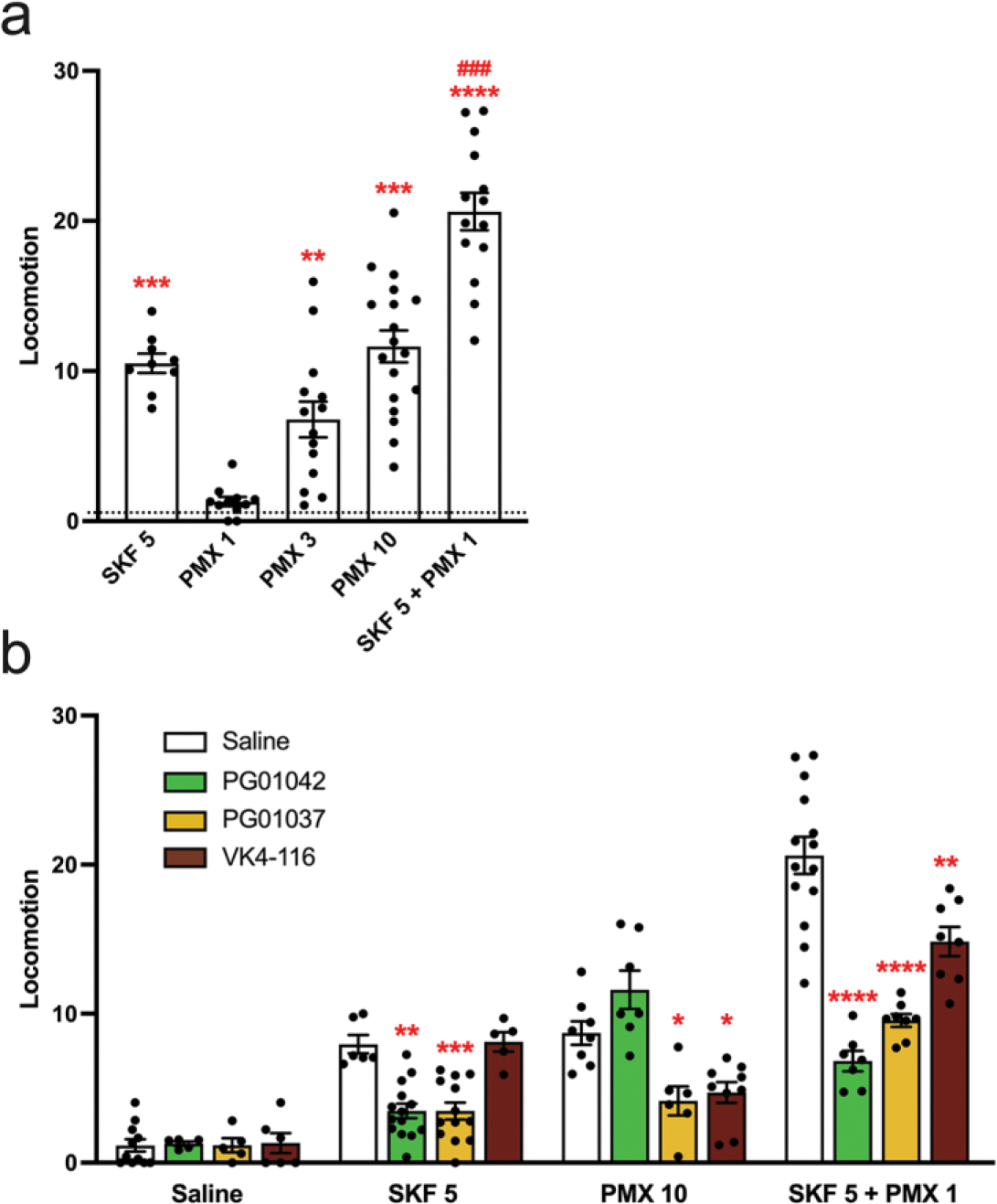
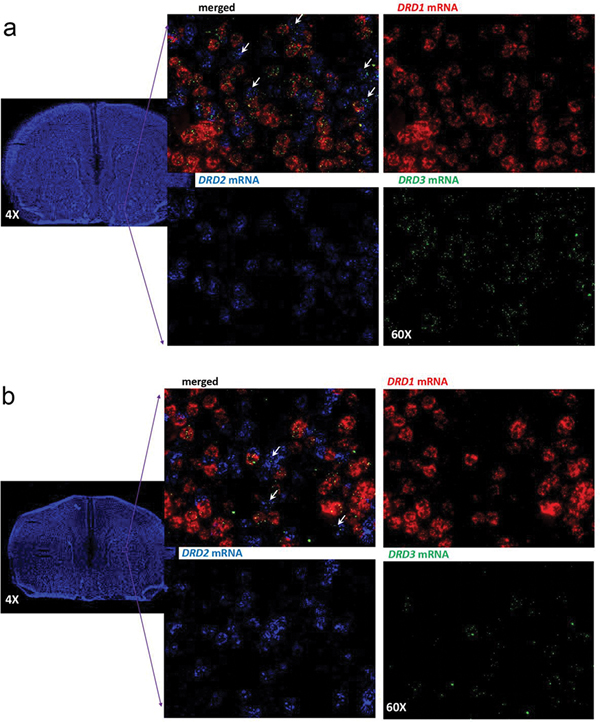
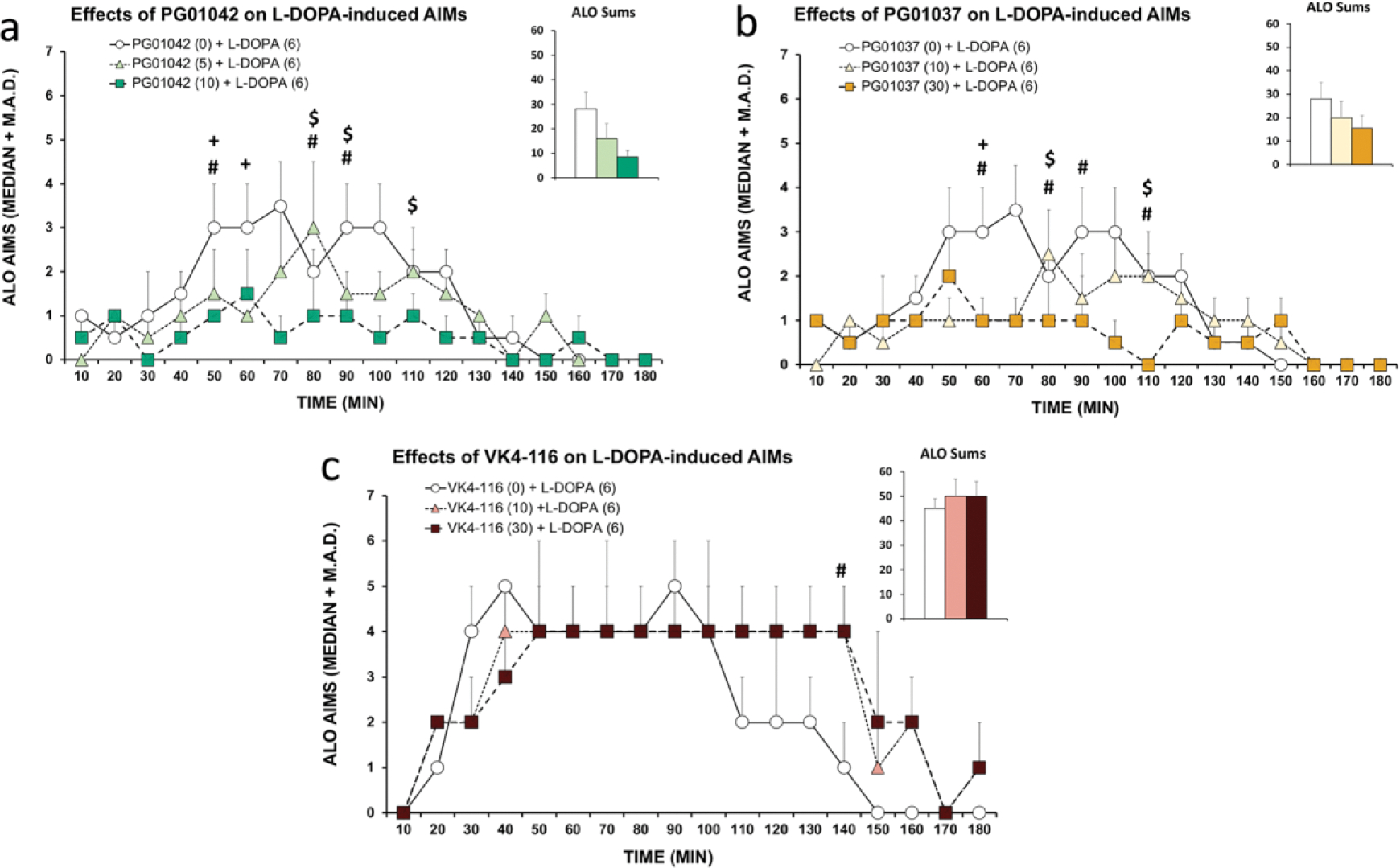
A main rationale for the role of G protein-coupled receptor (GPCR) heteromers as targets for drug development is the putative ability of selective ligands for specific GPCRs to change their pharmacological properties upon GPCR heteromerization. The present study provides a proof of concept for this rationale by demonstrating that heteromerization of dopamine D1 and D3 receptors (D1R and D3R) influences the pharmacological properties of three structurally similar selective dopamine D3R ligands, the phenylpiperazine derivatives PG01042, PG01037 and VK4-116. By using D1R-D3R heteromer-disrupting peptides, it could be demonstrated that the three D3R ligands display different D1R-D3R heteromer-dependent pharmacological properties: PG01042, acting as G protein-biased agonist, counteracted D1R-mediated signaling in the D1R-D3R heteromer; PG01037, acting as a D3R antagonist cross-antagonized D1R-mediated signaling in the D1R-D3R heteromer; and VK4-116 specifically acted as a ß-arrestin-biased agonist in the D1R-D3R heteromer. Molecular dynamics simulations predicted potential molecular mechanisms mediating these qualitatively different pharmacological properties of the selective D3R ligands that are dependent on D1R-D3R heteromerization. The results of in vitro experiments were paralleled by qualitatively different pharmacological properties of the D3R ligands in vivo. The results supported the involvement of D1R-D3R heteromers in the locomotor activation by D1R agonists in reserpinized mice and L-DOPA-induced dyskinesia in rats, highlighting the D1R-D3R heteromer as a main pharmacological target for L-DOPA-induced dyskinesia in Parkinson's disease. More generally, the present study implies that when suspecting its pathogenetic role, a GPCR heteromer, and not its individual GPCR units, should be considered as main target for drug development.
Keywords: Dopamine D(1) receptor; Dopamine D(3) receptor; G protein-coupled receptor (GPCR) heteromers; L-DOPA-induced dyskinesia; Locomotor activation; Mouse; PG01037 (PubChem CID: 11477180); PG01042 (PubChem CID: 11443078); Pramipexole (PubChem CID: 119570); Rat; SKF81297 (PubChem CID: 1218); VK4–116 (PubChem CID: 130431318).
Copyright © 2022. Published by Elsevier Ltd.
Conflict of interest statement
Declaration of Competing Interest The authors declare no competing interests.
Figures