

Primate hemorrhagic fever-causing arteriviruses are poised for spillover to humans
- PMID: 36182704
- PMCID: PMC9588614
- DOI: 10.1016/j.cell.2022.09.022
Primate hemorrhagic fever-causing arteriviruses are poised for spillover to humans
Abstract
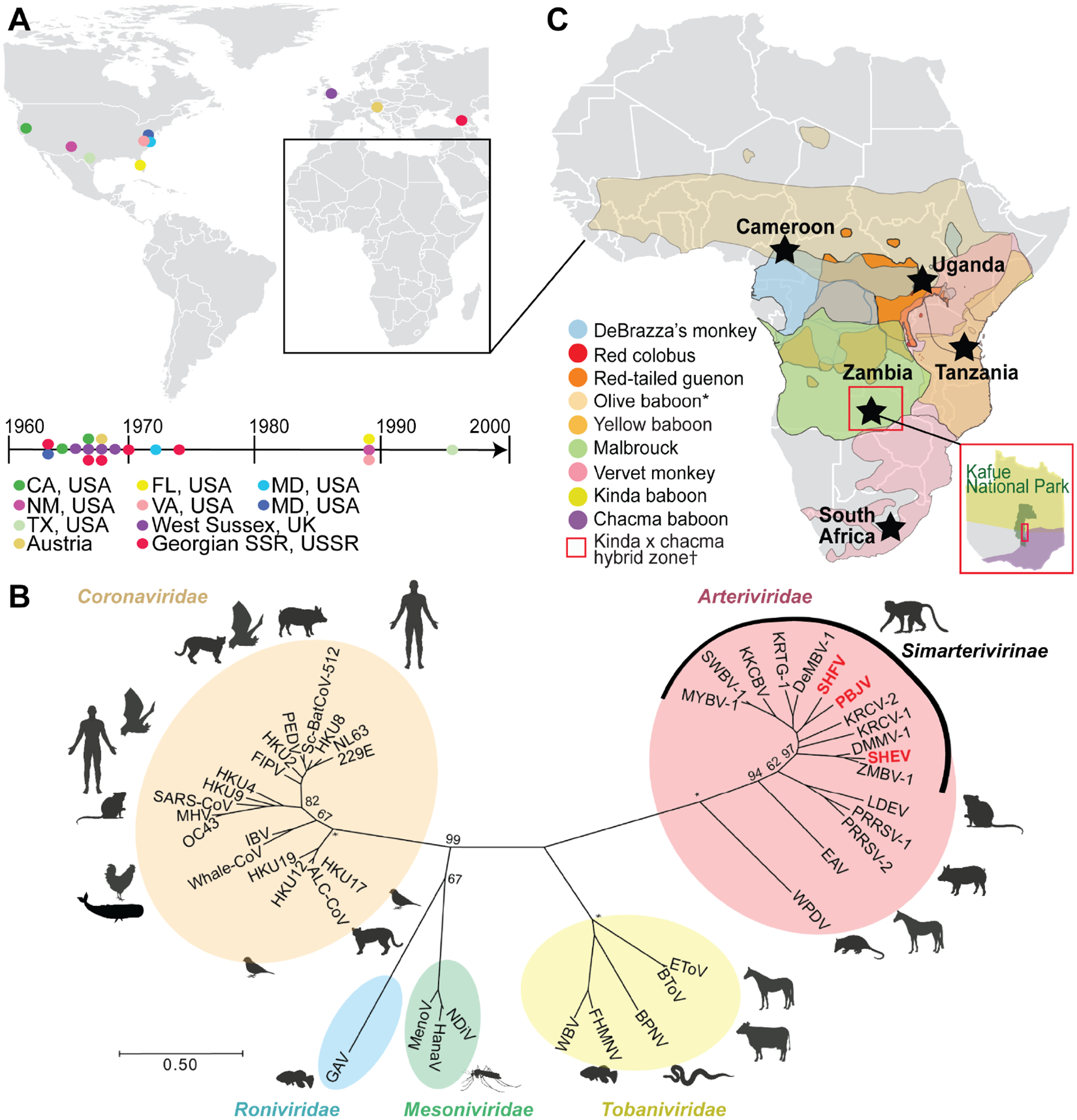
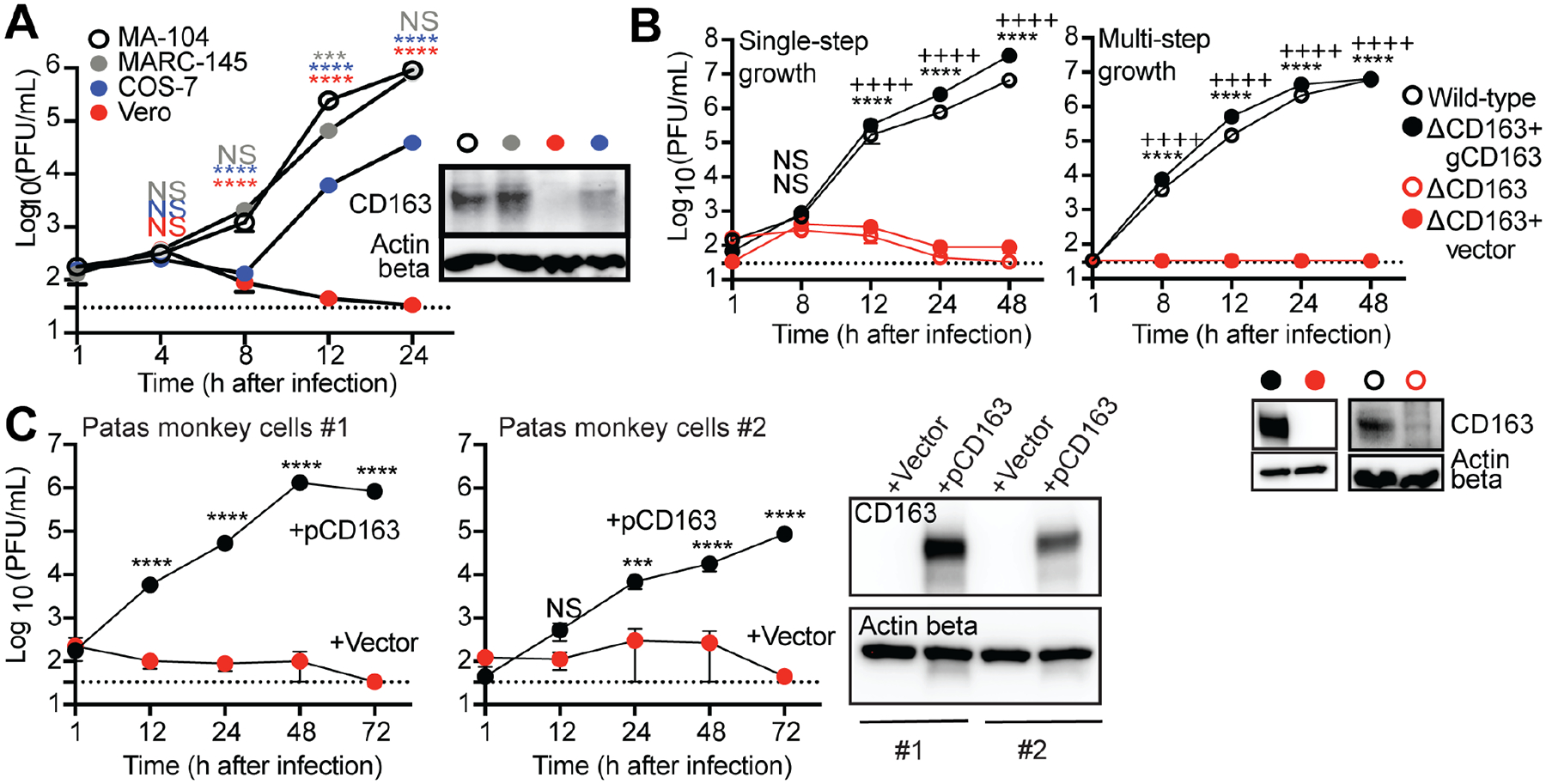
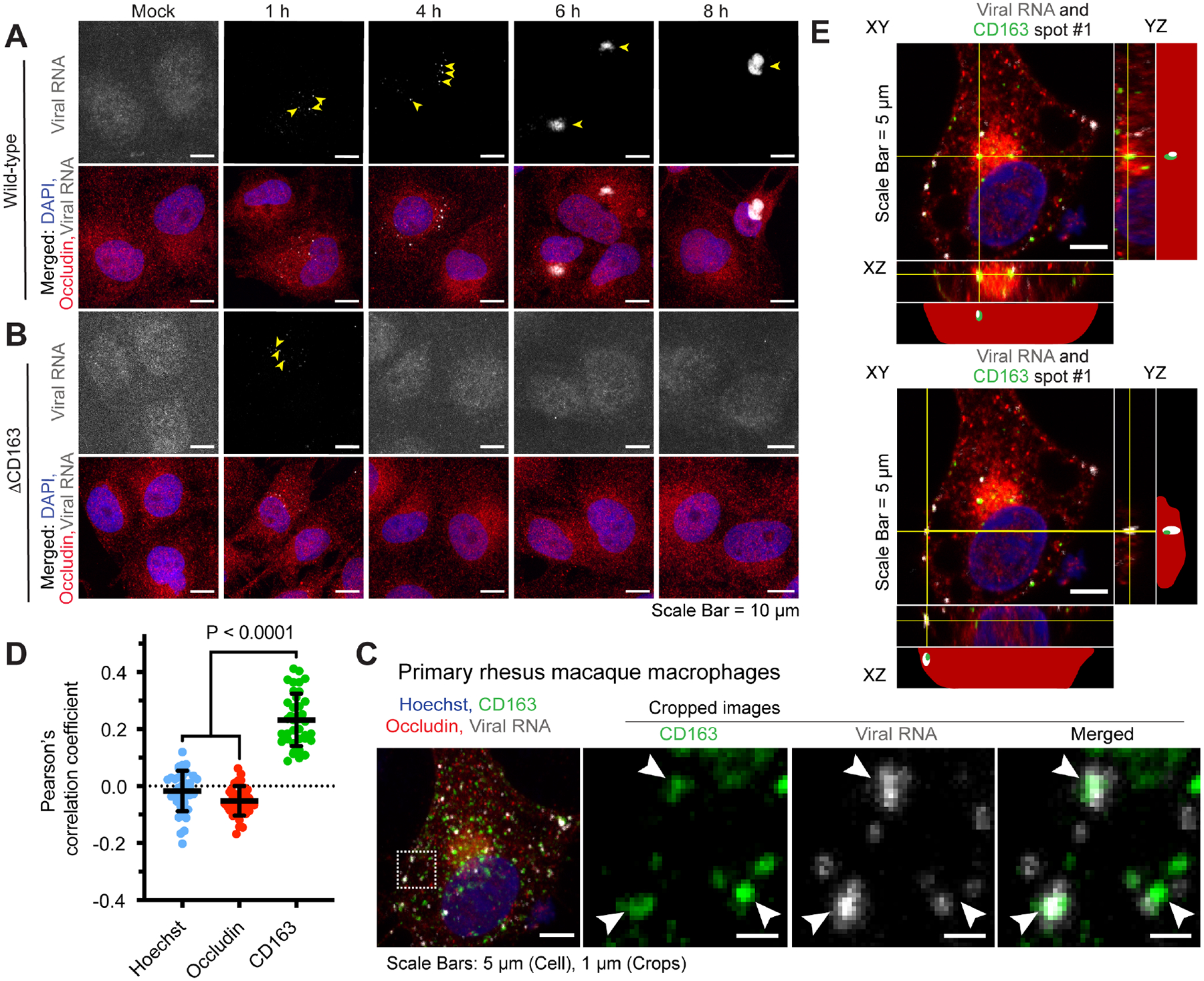
Simian arteriviruses are endemic in some African primates and can cause fatal hemorrhagic fevers when they cross into primate hosts of new species. We find that CD163 acts as an intracellular receptor for simian hemorrhagic fever virus (SHFV; a simian arterivirus), a rare mode of virus entry that is shared with other hemorrhagic fever-causing viruses (e.g., Ebola and Lassa viruses). Further, SHFV enters and replicates in human monocytes, indicating full functionality of all of the human cellular proteins required for viral replication. Thus, simian arteriviruses in nature may not require major adaptations to the human host. Given that at least three distinct simian arteriviruses have caused fatal infections in captive macaques after host-switching, and that humans are immunologically naive to this family of viruses, development of serology tests for human surveillance should be a priority.
Keywords: CD163 receptor; arms race; arteriviruses; disease emergence; hemorrhagic fever; positive selection; primates; virus entry; virus evolution; zoonosis.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.L.S. and Q.Y. are co-founders of, equity holders of, and consultants for Darwin Biosciences. S.L.S is on the scientific advisory board for Darwin Biosciences. S.L.S. serves as a consultant for the MITRE Corporation and is a member of the Planning Committee for Countering Zoonotic Spillover of High Consequence Pathogens, sponsored by the U.S. National Academies of Sciences, Engineering, and Medicine.
Figures
References
-
- Bailey AL, Lauck M, Weiler A, Sibley SD, Dinis JM, Bergman Z, Nelson CW, Correll M, Gleicher M, Hyeroba D, et al. (2014b). High genetic diversity and adaptive potential of two simian hemorrhagic fever viruses in a wild primate population. PLoS One 9, e90714. 10.1371/journal.pone.0090714. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
