

Leveraging network structure in nonlinear control
- PMID: 36182954
- PMCID: PMC9526710
- DOI: 10.1038/s41540-022-00249-2
Leveraging network structure in nonlinear control
Abstract
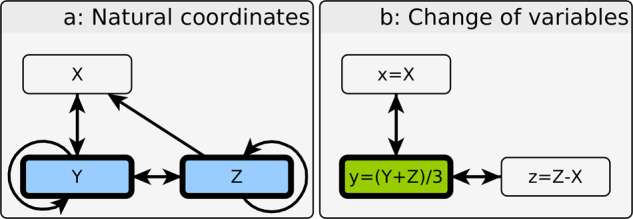
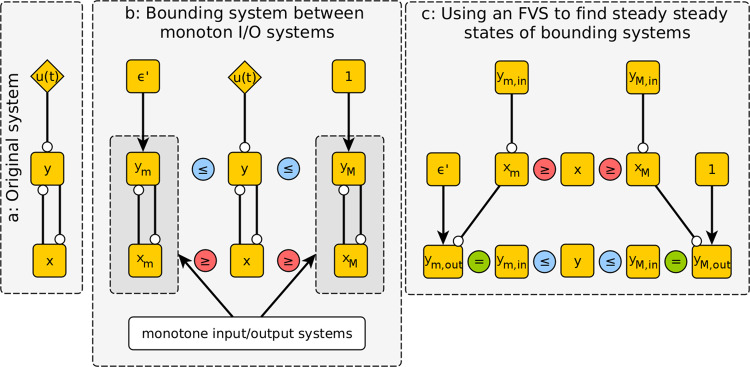
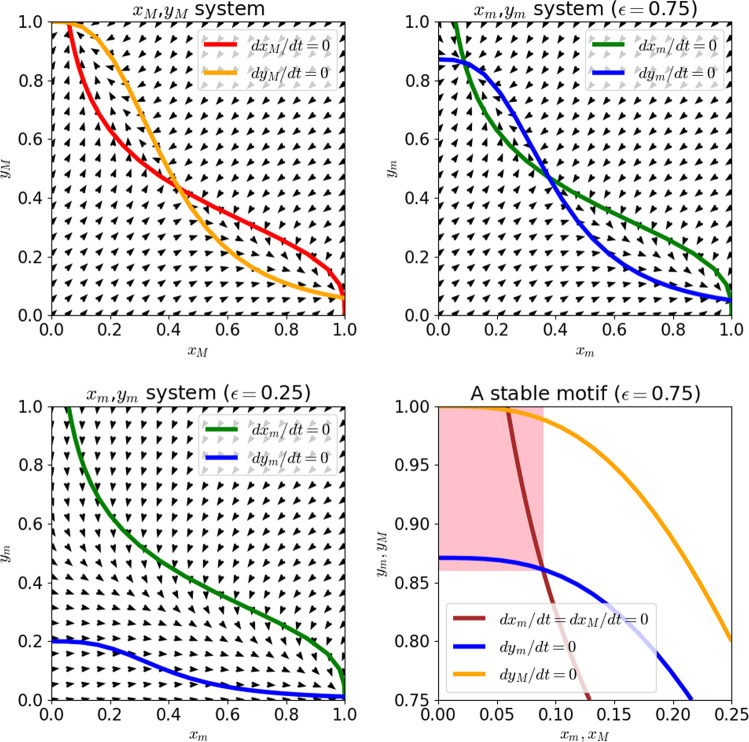
Over the last twenty years, dynamic modeling of biomolecular networks has exploded in popularity. Many of the classical tools for understanding dynamical systems are unwieldy in the highly nonlinear, poorly constrained, high-dimensional systems that often arise from these modeling efforts. Understanding complex biological systems is greatly facilitated by purpose-built methods that leverage common features of such models, such as local monotonicity, interaction graph sparsity, and sigmoidal kinetics. Here, we review methods for controlling the systems of ordinary differential equations used to model biomolecular networks. We focus on methods that make use of the structure of the network of interactions to help inform, which variables to target for control, and highlight the computational and experimental advantages of such approaches. We also discuss the importance of nonperturbative methods in biomedical and experimental molecular biology applications, where finely tuned interventions can be difficult to implement. It is well known that feedback loops, and positive feedback loops in particular, play a major determining role in the dynamics of biomolecular networks. In many of the methods we cover here, control over system trajectories is realized by overriding the behavior of key feedback loops.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
