

Regulation of factor V by the anticoagulant protease activated protein C: Influence of the B-domain and TFPIα
- PMID: 36183835
- PMCID: PMC9637641
- DOI: 10.1016/j.jbc.2022.102558
Regulation of factor V by the anticoagulant protease activated protein C: Influence of the B-domain and TFPIα
Abstract
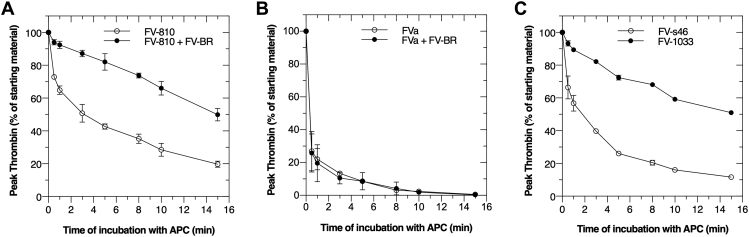
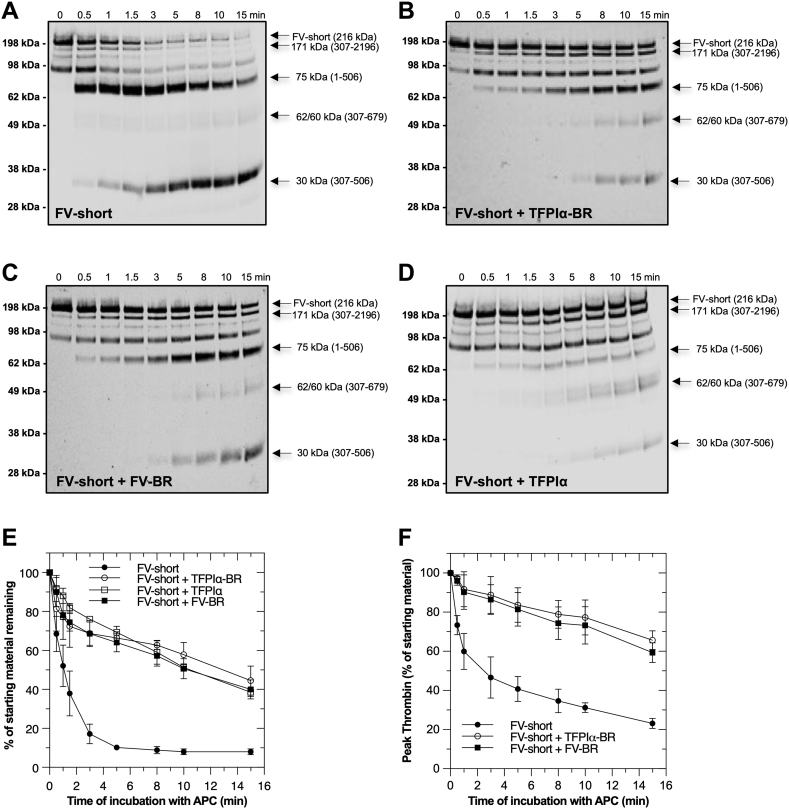
Activated protein C (APC) is an important anticoagulant protein that regulates thrombin generation through inactivation of factor V (FV) and activated factor V (FVa). The rate of APC inactivation of FV is slower compared to FVa, although proteolysis occurs at the same sites (Arg306, Arg506, and Arg679). The molecular basis for FV resistance to APC is unknown. Further, there is no information about how FV-short, a physiologically relevant isoform of FV with a shortened B-domain, is regulated by APC. Here, we identify the molecular determinants which differentially regulate APC recognition of FV versus FVa and uncover how FV-short can be protected from this anticoagulant pathway. Using recombinant FV derivatives and B-domain fragments, we show that the conserved basic region (BR; 963-1008) within the central portion of the B-domain plays a major role in limiting APC cleavage at Arg506. Derivatives of FV lacking the BR, including FV-short, were subject to rapid cleavage at Arg506 and were inactivated like FVa. The addition of a FV-BR fragment reversed this effect and delayed APC inactivation. We also found that anticoagulant glycoprotein TFPIα, which has a C-terminal BR homologous to FV-BR, protects FV-short from APC inactivation by delaying cleavage at Arg506. We conclude that the FV-BR plays a major role in protecting FV from APC inactivation. Using a similar mechanistic strategy, TFPIα also shields FV-short from APC. These findings clarify the resistance of FV to APC, advance our understanding of FV/FVa regulation, and establish a mechanistic framework for manipulating this reaction to alter coagulation.
Keywords: acidic and basic residues; coagulation factor; cofactor; factor V; factor Va; hemostasis; procofactor; protein complex; prothrombinase; thrombin; tissue factor pathway inhibitor.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures




References
-
- Mann K.G., Nesheim M.E., Church W.R., Haley P.E., Krishnaswamy S. Surface dependent reactions of the vitamin K-dependent enzyme complexes. Blood. 1990;76:1–16. - PubMed
-
- Nesheim M.E., Taswell J.B., Mann K.G. The contribution of bovine factor V and factor Va to the activity of prothrombinase. J. Biol. Chem. 1979;254:10952–10962. - PubMed
-
- Toso R., Camire R.M. Removal of B-domain sequences from factor V rather than specific proteolysis underlies the mechanism by which cofactor function is realized. J. Biol. Chem. 2004;279:21643–21650. - PubMed
-
- Zhu H., Toso R., Camire R.M. Inhibitory sequences within the B-domain stabilize circulating factor V in an inactive state. J. Biol. Chem. 2007;282:15033–15039. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
