

Dysregulated DNA methylation in the pathogenesis of Fabry disease
- PMID: 36186841
- PMCID: PMC9519376
- DOI: 10.1016/j.ymgmr.2022.100919
Dysregulated DNA methylation in the pathogenesis of Fabry disease
Abstract
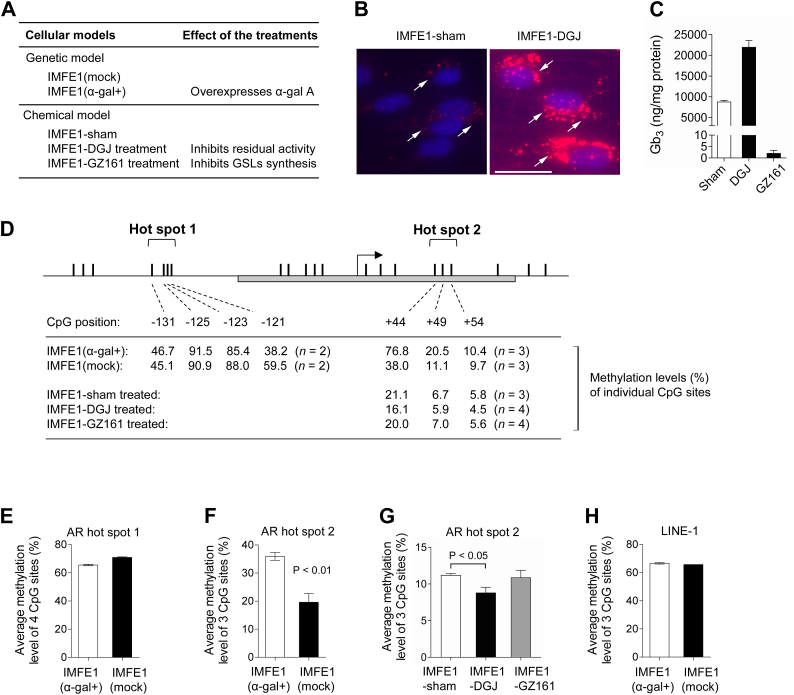
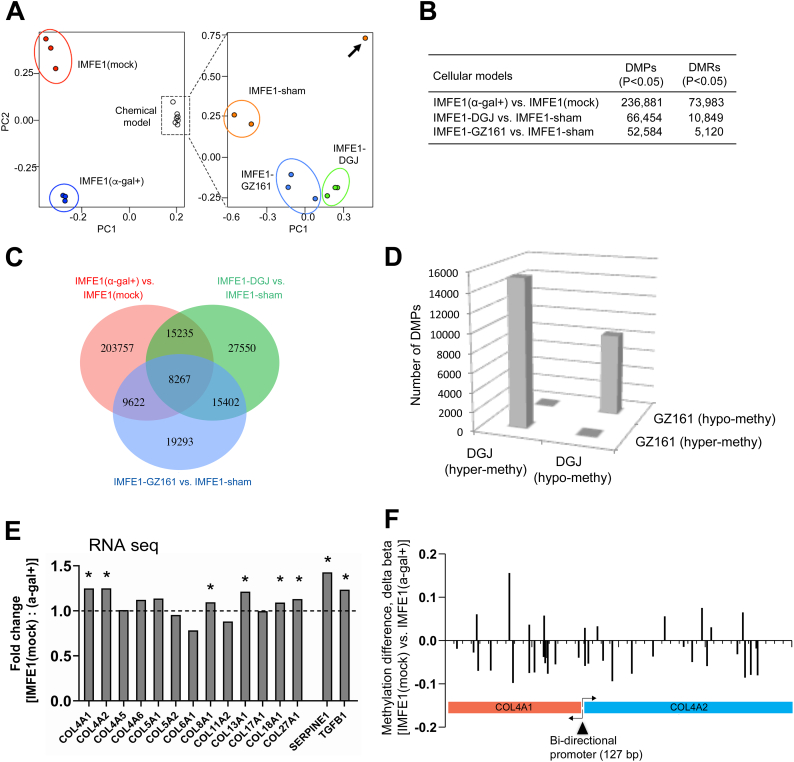
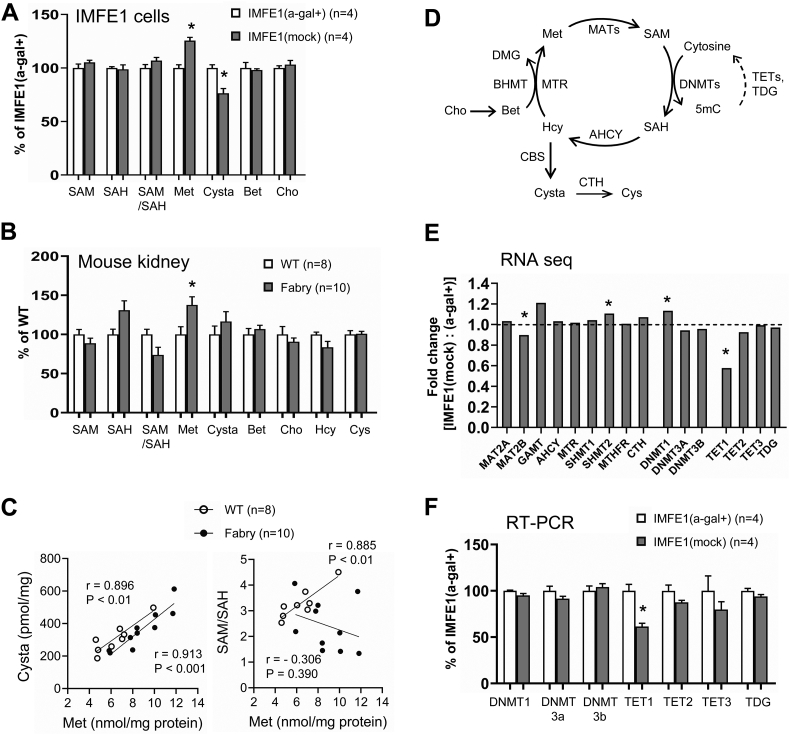
Fabry disease is an X-linked lysosomal storage disorder caused by a deficiency of α-galactosidase A and subsequent accumulation of glycosphingolipids with terminal α-D-galactosyl residues. The molecular process through which this abnormal metabolism of glycosphingolipids causes multisystem dysfunction in Fabry disease is not fully understood. We sought to determine whether dysregulated DNA methylation plays a role in the development of this disease. In the present study, using isogenic cellular models derived from Fabry patient endothelial cells, we tested whether manipulation of α-galactosidase A activity and glycosphingolipid metabolism affects DNA methylation. Bisulfite pyrosequencing revealed that changes in α-galactosidase A activity were associated with significantly altered DNA methylation in the androgen receptor promoter, and this effect was highly CpG loci-specific. Methylation array studies showed that α-galactosidase A activity and glycosphingolipid levels were associated with differential methylation of numerous CpG sites throughout the genome. We identified 15 signaling pathways that may be susceptible to methylation alterations in Fabry disease. By incorporating RNA sequencing data, we identified 21 genes that have both differential mRNA expression and methylation. Upregulated expression of collagen type IV alpha 1 and alpha 2 genes correlated with decreased methylation of these two genes. Methionine levels were elevated in Fabry patient cells and Fabry mouse tissues, suggesting that a perturbed methionine cycle contributes to the observed dysregulated methylation patterns. In conclusion, this study provides evidence that α-galactosidase A deficiency and glycosphingolipid storage may affect DNA methylation homeostasis and highlights the importance of epigenetics in the pathogenesis of Fabry disease and, possibly, of other lysosomal storage disorders.
Keywords: DNA methylation; Deoxygalactonojirimycin; Fabry disease; Globotriaosylceramide; Substrate reduction therapy; α-Galactosidase A.
© 2022 Published by Elsevier Inc.
Conflict of interest statement
None.
Figures
References
-
- Brady R., Gal A.E., Bradley R.M., Martensson E., Warshaw A.L., Laster L. Enzymatic defect in Fabry disease: ceramide trihexosidase deficiency. N. Engl. J. Med. 1967;276:1163–1167. - PubMed
-
- Desnick R.J., Ioannou Y.A., Eng C.M. In: In The Metabolic and Molecular Bases of Inherited Disease. Scriver C.R., Beaudet A.L., Sly W.S., Valle D., editors. McGraw-Hill; New York: 2001. pp. 3733–3774.
-
- Schiffmann R. Fabry disease. Pharmacol. Ther. 2009;122:65–77. - PubMed
-
- Moore D.F., Gelderman M.P., Ferreira P.A., Fuhrmann S.R., Yi H., Elkahloun A., Lix L.M., Brady R.O., Schiffmann R., Goldin E. Genomic abnormalities of the murine model of Fabry disease after disease-related perturbation, a systems biology approach. Proc. Natl. Acad. Sci. U. S. A. 2007;104:8065–8070. - PMC - PubMed
LinkOut - more resources
Full Text Sources
