

Piezo buffers mechanical stress via modulation of intracellular Ca2+ handling in the Drosophila heart
- PMID: 36187790
- PMCID: PMC9515499
- DOI: 10.3389/fphys.2022.1003999
Piezo buffers mechanical stress via modulation of intracellular Ca2+ handling in the Drosophila heart
Abstract
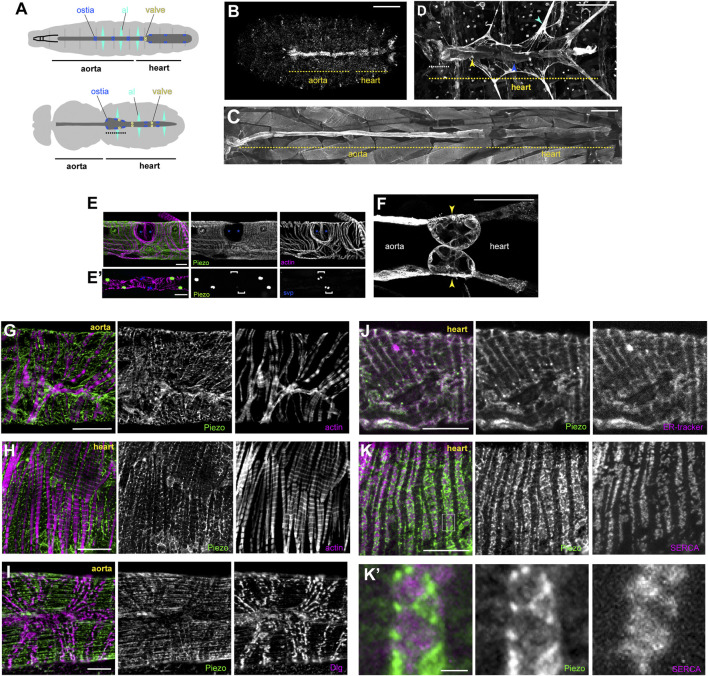
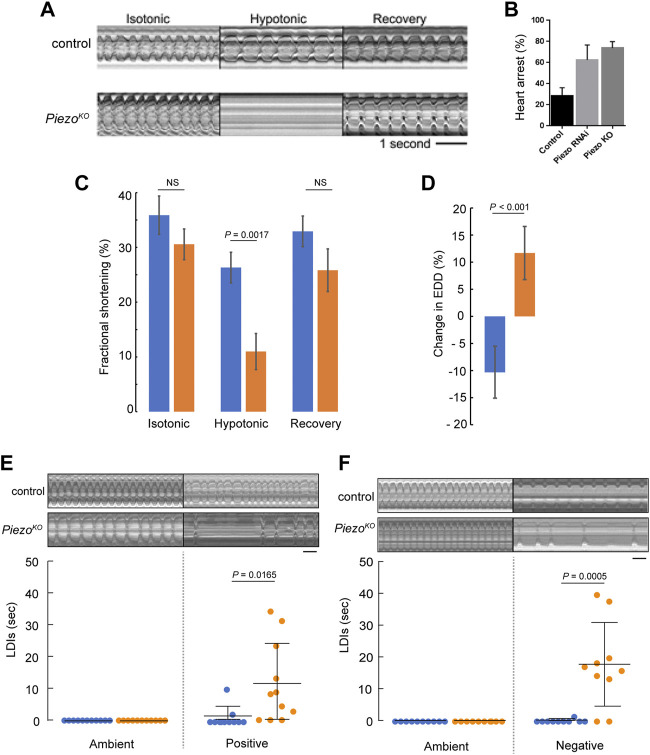
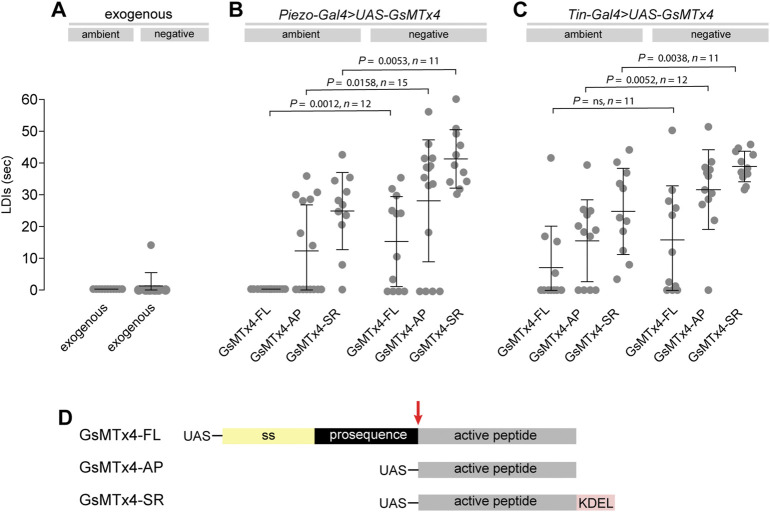
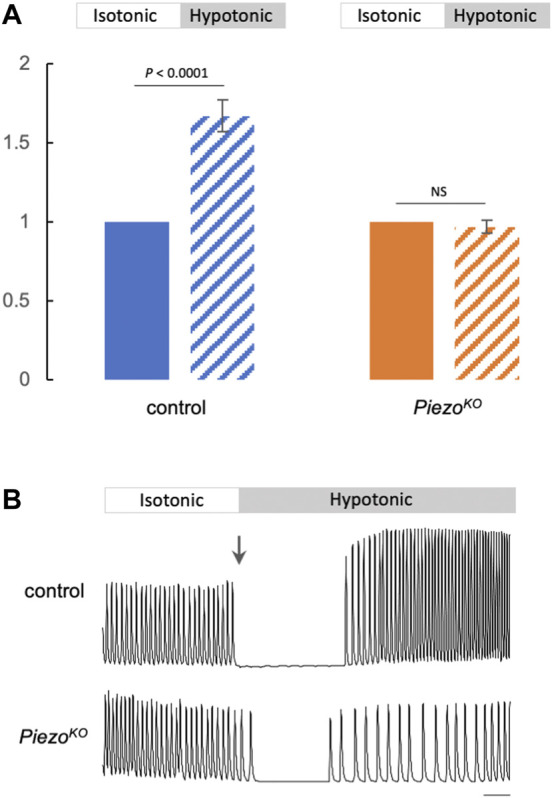
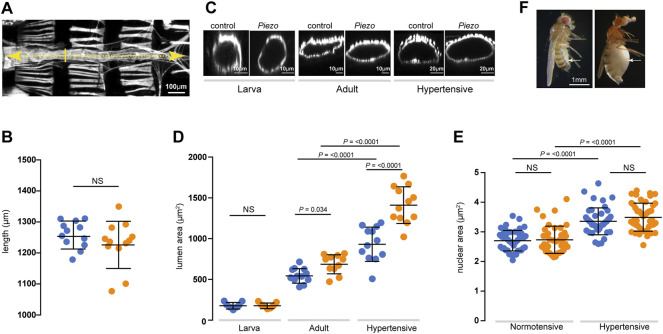
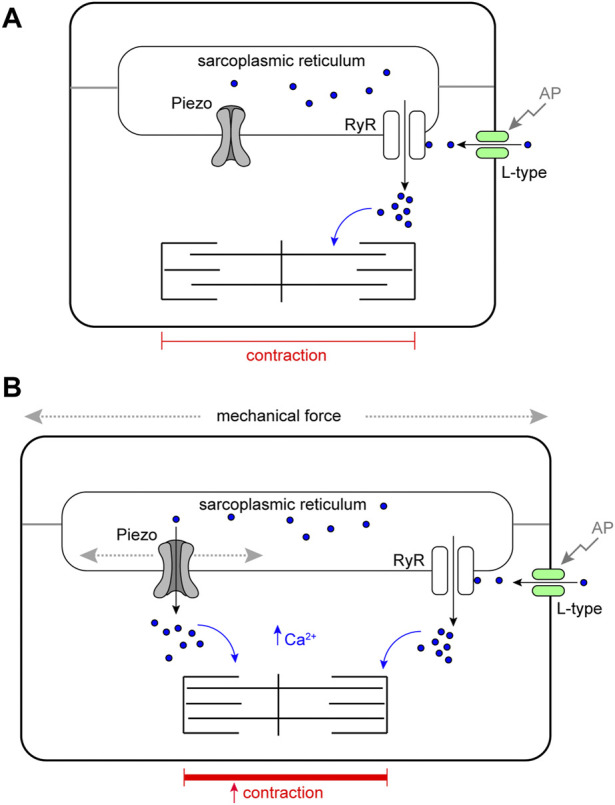
Throughout its lifetime the heart is buffeted continuously by dynamic mechanical forces resulting from contraction of the heart muscle itself and fluctuations in haemodynamic load and pressure. These forces are in flux on a beat-by-beat basis, resulting from changes in posture, physical activity or emotional state, and over longer timescales due to altered physiology (e.g. pregnancy) or as a consequence of ageing or disease (e.g. hypertension). It has been known for over a century of the heart's ability to sense differences in haemodynamic load and adjust contractile force accordingly (Frank, Z. biology, 1895, 32, 370-447; Anrep, J. Physiol., 1912, 45 (5), 307-317; Patterson and Starling, J. Physiol., 1914, 48 (5), 357-79; Starling, The law of the heart (Linacre Lecture, given at Cambridge, 1915), 1918). These adaptive behaviours are important for cardiovascular homeostasis, but the mechanism(s) underpinning them are incompletely understood. Here we present evidence that the mechanically-activated ion channel, Piezo, is an important component of the Drosophila heart's ability to adapt to mechanical force. We find Piezo is a sarcoplasmic reticulum (SR)-resident channel and is part of a mechanism that regulates Ca2+ handling in cardiomyocytes in response to mechanical stress. Our data support a simple model in which Drosophila Piezo transduces mechanical force such as stretch into a Ca2+ signal, originating from the SR, that modulates cardiomyocyte contraction. We show that Piezo mutant hearts fail to buffer mechanical stress, have altered Ca2+ handling, become prone to arrhythmias and undergo pathological remodelling.
Keywords: Drosophila; Frank-Starling; Piezo; calcium; heart; mechanosensitive ion channel; mechanotransduction; sacroplasmic recticulum.
Copyright © 2022 Zechini, Camilleri-Brennan, Walsh, Beaven, Moran, Hartley, Diaz and Denholm.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous