

Structural and Mechanistic Bases of Viral Resistance to HIV-1 Capsid Inhibitor Lenacapavir
- PMID: 36190128
- PMCID: PMC9600929
- DOI: 10.1128/mbio.01804-22
Structural and Mechanistic Bases of Viral Resistance to HIV-1 Capsid Inhibitor Lenacapavir
Abstract
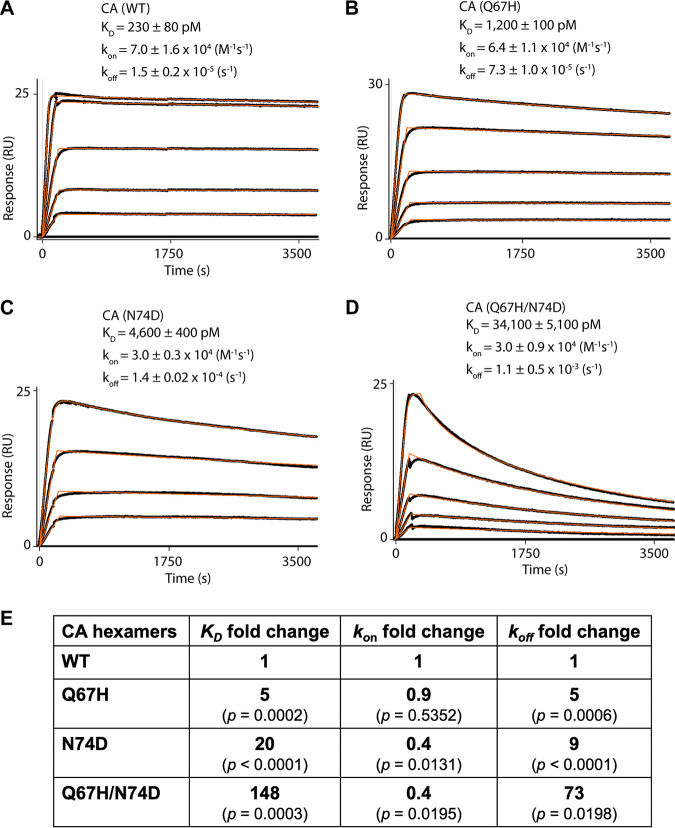
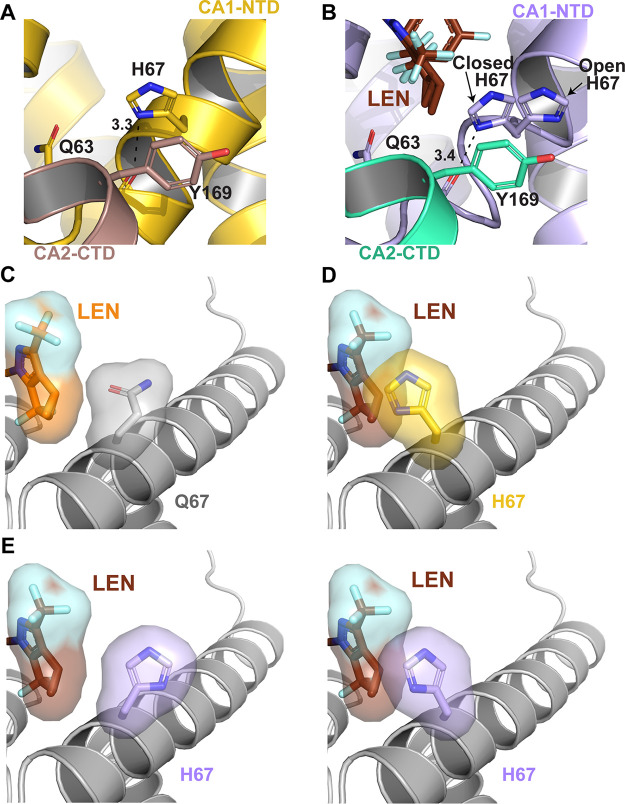
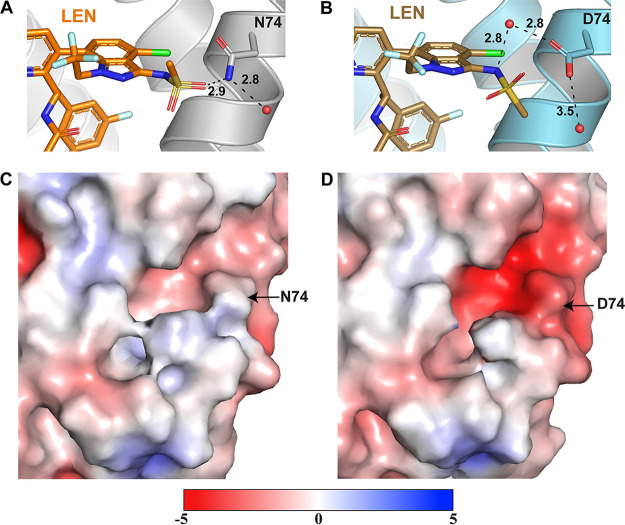
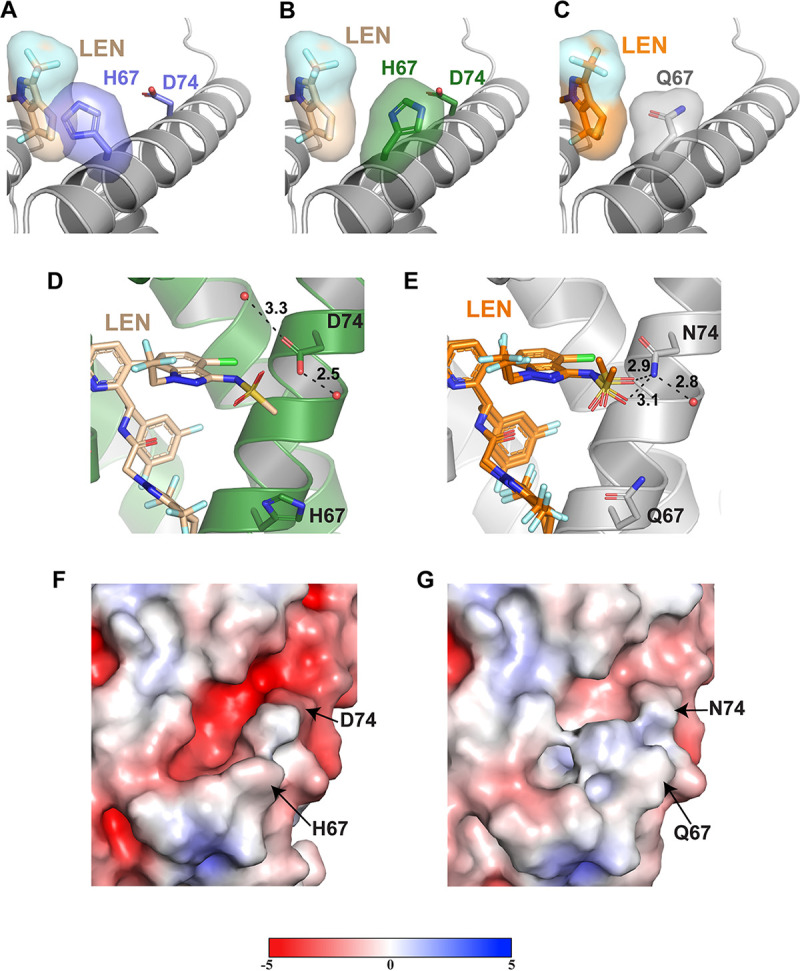
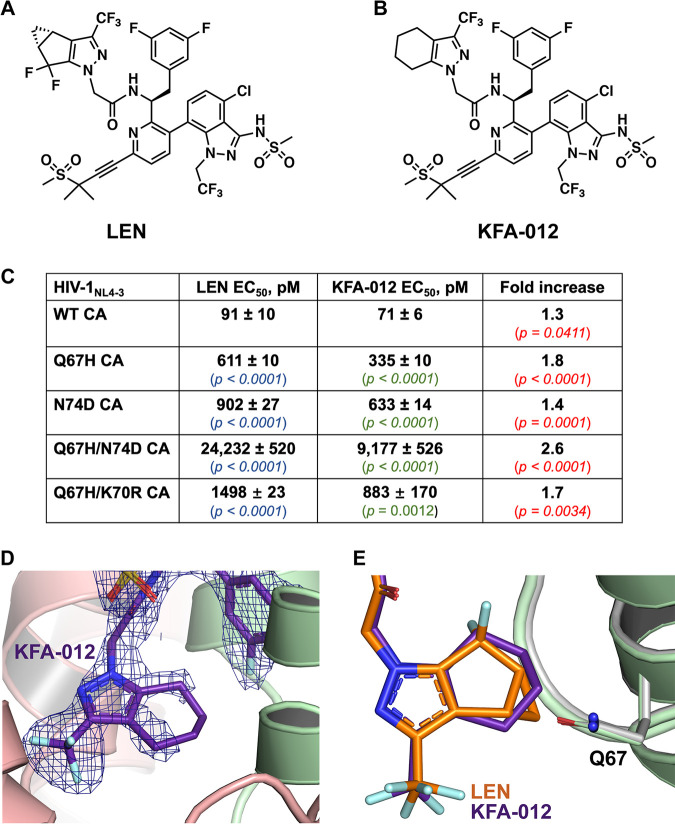
Lenacapavir (LEN) is a long-acting, highly potent HIV-1 capsid (CA) inhibitor. The evolution of viral variants under the genetic pressure of LEN identified Q67H, N74D, and Q67H/N74D CA substitutions as the main resistance associated mutations (RAMs). Here, we determined high-resolution structures of CA hexamers containing these RAMs in the absence and presence of LEN. Our findings reveal that the Q67H change induces a conformational switch, which adversely affects the inhibitor binding. In the unliganded protein, the His67 side chain adopts the closed conformation by projecting into the inhibitor binding pocket and thereby creating steric hindrance with respect to LEN. Upon the inhibitor binding, the His67 side chain repositions to the open conformation that closely resembles the Gln67 side chain in the WT protein. We propose that the switch from the closed conformation to the open conformation, which is needed to accommodate LEN, accounts for the reduced inhibitor potency with respect to the Q67H CA variant. The N74D CA change results in the loss of a direct hydrogen bond and in induced electrostatic repulsions between CA and LEN. The double Q67H/N74D substitutions exhibited cumulative effects of respective single amino acid changes. An examination of LEN binding kinetics to CA hexamers revealed that Q67H and N74D CA changes adversely influenced the inhibitor binding affinity (KD) by primarily affecting the dissociation rate constant (koff). We used these structural and mechanistic findings to rationally modify LEN. The resulting analog exhibited increased potency against the Q67H/N74D viral variant. Thus, our studies provide a means for the development of second-generation inhibitors with enhanced barriers to resistance. IMPORTANCE LEN is an investigational long-acting agent for future HIV-1 treatment regimens. While ongoing clinical trials have highlighted a largely beneficial profile of LEN for the treatment of HIV-1 infected people with limited therapy options, one notable shortcoming is a relatively low barrier of viral resistance to the inhibitor. Cell culture-based viral breakthrough assays identified N74D, Q67H, and N74D/Q67H capsid changes as the main resistance associated mutations (RAMs). N74D and Q67H capsid substitutions have also emerged in clinical trials in some patients who received subcutaneous LEN. Understanding the structural basis behind viral resistance to LEN is expected to aid in the rational development of improved inhibitors with enhanced barriers to resistance. Here, we report high resolution structures of the main drug resistant capsid variants, which provide mechanistic insight into the viral resistance to LEN. We used these findings to develop an improved inhibitor, which exhibited enhanced activity against the viral Q67H/N74D capsid phenotype compared with that of parental LEN.
Keywords: HIV-1; antiretroviral agents; capsid; drug resistance mechanisms; structure.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Link JO, Rhee MS, Tse WC, Zheng J, Somoza JR, Rowe W, Begley R, Chiu A, Mulato A, Hansen D, Singer E, Tsai LK, Bam RA, Chou CH, Canales E, Brizgys G, Zhang JR, Li J, Graupe M, Morganelli P, Liu Q, Wu Q, Halcomb RL, Saito RD, Schroeder SD, Lazerwith SE, Bondy S, Jin D, Hung M, Novikov N, Liu X, Villasenor AG, Cannizzaro CE, Hu EY, Anderson RL, Appleby TC, Lu B, Mwangi J, Liclican A, Niedziela-Majka A, Papalia GA, Wong MH, Leavitt SA, Xu Y, Koditek D, Stepan GJ, Yu H, Pagratis N, Clancy S, Ahmadyar S, et al. 2020. Clinical targeting of HIV capsid protein with a long-acting small molecule. Nature 584:614–618. doi: 10.1038/s41586-020-2443-1. - DOI - PMC - PubMed
-
- Yant SR, Mulato A, Hansen D, Tse WC, Niedziela-Majka A, Zhang JR, Stepan GJ, Jin D, Wong MH, Perreira JM, Singer E, Papalia GA, Hu EY, Zheng J, Lu B, Schroeder SD, Chou K, Ahmadyar S, Liclican A, Yu H, Novikov N, Paoli E, Gonik D, Ram RR, Hung M, McDougall WM, Brass AL, Sundquist WI, Cihlar T, Link JO. 2019. A highly potent long-acting small-molecule HIV-1 capsid inhibitor with efficacy in a humanized mouse model. Nat Med 25:1377–1384. doi: 10.1038/s41591-019-0560-x. - DOI - PMC - PubMed
-
- Bester SM, Wei G, Zhao H, Adu-Ampratwum D, Iqbal N, Courouble VV, Francis AC, Annamalai AS, Singh PK, Shkriabai N, Van Blerkom P, Morrison J, Poeschla EM, Engelman AN, Melikyan GB, Griffin PR, Fuchs JR, Asturias FJ, Kvaratskhelia M. 2020. Structural and mechanistic bases for a potent HIV-1 capsid inhibitor. Science 370:360–364. doi: 10.1126/science.abb4808. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
